Chapter VII. Coelenterata. The Sub-Kingdom Coelenterata
Description
This section is from the "" book, by .
Chapter VII. Coelenterata. The Sub-Kingdom Coelenterata
1. Characters of the Sub-Kingdom. 2. Divisions. 3. General Characters of the Hydrozoa. 4. Explanation of Technical Terms.
The Sub-kingdom Coelenterata (Frey and Leuckhart) may be considered as a modern representative of the Radiata of Cuvier. From the Radiata, however, the Echinodermata and Rotifera have been removed, the entire sub-kingdom of the Protozoa has been taken away, and the Polyzoa have been relegated to their proper place amongst the Mollusca. Deducting these groups from the old Radiata, the residue, comprising most of the animals commonly known as Polypes or Zoophytes, remains to constitute the modern Coelenterata.
The Coelenterata may be defined as animals whose alimentary canal communicates freely with the general cavity of the body ("somatic cavity"). The substance of the body is made up of t7vo fundamental membranes - an outer layer, called the "ectoderm" and an inner layer, or "endoderm." There are no distinct neural and hoemal regions, and in the great majority of the members of the sub-kingdom there are no traces of a nervous system. Peculiar uriicating organs, or "thread-cells," are usually present; and, generally speaking, a radiate condition of the organs is perceptible, especially in the tentacles with which most are provided. In all the Coelenterata distinct reproductive organs have been shown to exist.
The leading feature which distinguishes the Coelenterata, and the one from which the name of the sub-kingdom is derived, is the peculiar structure of the digestive system. In the Protozoa, as we have seen, a mouth is only present in the higher forms, and in no case is there any definite internal cavity bounded by the walls of the body to which the name of "body-cavity" or "somatic cavity" could be properly applied (unless it be allowed that such really exists in the sponges). In animals higher than the Coelenterata, on the other hand, there is not only generally a permanent mouth, but the walls of the body usually enclose a permanent chamber or "body-cavity." Further, in most cases, the mouth conducts into an alimentary canal, which is always distinct from the body-cavity, never opening into it, but usually passing through it to open on the surface by another distinct aperture (the anus). In most cases, therefore, the alimentary canal is a tube which communicates with the outer world by two apertures - a mouth and anus - but which simply passes through the body-cavity without in any way communicating with it. In the Coelenterata (fig. 35) there is an intermediate condition of parts. There is a distinct and permanent mouth, and a distinct and permanent body-cavity, but the mouth opens into, and communicates freely with, the body-cavity. In some cases (Hydrozoa) the mouth opens directly into the general body-cavity, which then serves as a digestive cavity as well (fig. 37). In other cases there intervenes between the mouth and the body-cavity a short alimentary tube, which communicates externally with the outer world through the mouth, and opens below by a wide aperture into the general cavity of the body (Actinozoa, fig. 35). In no case is there a distinct intestinal canal which runs through the body and opens on the surface by a mouth at one end and an excretory aperture or anus at the other. It should, however, be mentioned here that some modern zoologists, such as Gegen-baur and Haeckel, consider that the entire system of internal cavities in any Coelenterate is truly homologous with the intestinal canal of other animals, and that the Coelenterata, therefore, possess no true body-cavity at all. To this view some of our most distinguished authorities, such as Professor Allman, have given their adhesion; and there is no doubt that there are weighty grounds for regarding it as the correct explanation of the facts, though any discussion of these grounds would be out of place here. It need only be added that if this view be accepted, it will entirely subvert the generally received conception of the structure of the Coelenterata as above expressed.

Fig. 35. - Diagrammatic vertical section of a Sea-Anemone. a Mouth; .s Stomach; b Body-cavity ; c c Convoluted cords ("craspeda") containing thread-cells, and forming the free edges of the mesentery (m); t, t Tentacles ; o Reproductive organ contained within the mesentery. The ectoderm (e) is indicated by the broad external line, the endoderm (e') by the thin line and the space between that and the ectoderm.
Though of the true "radiate" type, some Coelenterates show traces of bilateral symmetry. Thus, in some Sea-anemones one of the tentacles is larger than, or differently coloured from, the others; and in some corals two of the primary septa, opposite one another, are larger than the rest, and divide the animal into two halves.
With regard to the fundamental tissues of the Coelenterata, there exist two primary membranes, of which one forms the outer surface of the body, and is called the "ectoderm;" whilst the other lines the alimentary canal, the general cavity of the body, and the tubular tentacles, and is termed the "endoderm." These membranes correspond with the primitive serous and mucous layers ("epiblast" and "hypoblast") of the germinal area, and become differentiated in opposite directions, the ectoderm growing from within outwards, the endoderm from without inwards. Each is primitively cellular in its minute structure, and each may be rendered more or less complex by vacuolation or fibrillation. Between the ectoderm and endoderm there is sometimes a third layer ("mesoderm" or "mesoblast"), which is commonly of a muscular nature.
In connection with the integument of the Coelenterata, the organs termed "thread-cells" ("cnidae," or "nematocysts") must be noticed. These are peculiar cellular bodies (fig. 36), of various shapes, which probably serve as weapons of offence and defence, and which communicate to many members of the sub-kingdom (e.g., the Sea-blubbers) their well-known power of stinging. In the common Hydra the thread-cells (fig. 36, E) consist of "oval elastic sacs, containing a long coiled filament, barbed at its base, and serrated along its edges. When fully developed the sacs are tensely filled with fluid, and the slightest touch is sufficient to cause the retroversion of the filament, which then projects beyond the sac for a distance, which is not uncommonly equal to many times the length of the latter" * (Huxley). Many beautiful modifications of shape are known in the thread-cells of different Coelenterates, but their essential structure in all cases is much the same as in the Hydra. It is only in few cases, comparatively speaking, that the thread-cells have the power of piercing and irritating the human skin ; but even in the diminutive Hydra it is probable that they exercise some benumbing and deleterious influence on the living organisms which may be captured as prey. Besides the thread-cells, the tentacles of some Hydroids are furnished with rigid hair-like processes, which are probably tactile in function, and which are known as "palpocils."
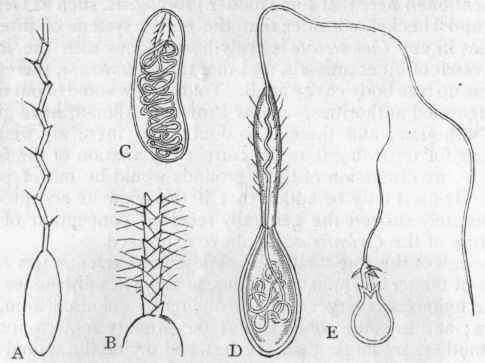
Fig. 36. - Thread-cells of Coelenterata, greatly magnified. A and B, The thread-cell of Caryophyllia Smithii, in the everted condition, and in two varieties ; C and D, The thread-cell of Corallimorphus profundus, in a quiescent and active condition, enlarged about 500 times; E, The thread-cell of Hydra, in an everted condition. (After Gosse and Moseley.)
The Coelenterata are divided into two classes, termed respectively the Hydrozoa and the Actinozoa.
* Thread-cells, though very commonly, if not universally, present in the Coelenterata, are nevertheless not peculiar to them. Similar organs have been shown to exist in several of the Nudibranchiate Mollusca, as well as in some Annelides (Spio seticornis). There likewise exist analogous organs trichocysts) in several of the Infusoria, and in the Planarida.
Continue to:
My Books

