Annelida. Part 18
Description
This section is from the book "General Outline Of The Organization Of The Animal Kingdom, And Manual Of Comparative Anatomy", by Thomas Rymer Jones. Also available from Amazon: A General Outline of the Animal Kingdom and Manual of Comparative Anatomy.
Annelida. Part 18
Each branchial tuft and each individual vessel possesses an independent power of contraction: in the contracted state the tuft almost disappears, so completely effected is the emptying of the vessels. The contraction or systole in any given tuft occurs at frequent but irregular intervals: this movement does not take place simultaneously in all the branchiae, but at different periods in different tufts. The vessels have the appearance of being quite naked; and if examined in the living state, each ramuscule seems to consist of only a single trunklet: if this were really the case, it would of course resolve itself into a tube ending in a cul-de-sac, and the blood-movement would be a flux and reflux; but by injection it is easy to show that the finest division of the branchial arbusculc contains a double vessel enveloped in a common muscular (although extremely diaphanous) sheath. That these vascular sheaths, which are only fine productions of the integuments, are furnished with voluntary muscular fibres, is proved by the rapid and simultaneous retraction of all the branchiae into the interior of the body which follows when the animal is touched.
In Arenicola, as in all Annelida in which the vessels of these organs are naked, the branchiae are destitute of vibratile cilia; and it will be found that under such circumstances, namely when the branchial vessels occur as naked projections from the external surface, the description here given of these organs in Arenicola will apply in every respect to all other Annelida so furnished. It will be observed that in all the dorsibranchiate genera furnished with branchiae such as those described above, the true blood, circulating in its proper vessels, is found to be exclusively the seat and subject of the respiratory process; the fluid in the peritoneal cavity, abundant in quantity and highly organized though it be, does not in the least degree participate in this great function. The Dorsibranchiate Annelids may, however, be divided into two great groups, of which one would comprehend those genera in which the function of breathing devolves exclusively on the true blood, while the other would be characterized by the fact that the branchiae are constructed so as to permit more or less completely the exposure, in conjunction with the blood-proper, of the chylaqueous fluid of the visceral cavity to the influence of the surrounding aerating element.
Thus it will be seen that when the branchial apparatus is penetrated by two separate and distinct fluids, coordinate probably in organic properties, the vascular system of the body generally will be found so much the less developed, in proportion as the peritoneal fluid supplants the blood in the branchiae. In those races of Dorsibranchiate worms possessing both these kinds of circulation, naked unciliated blood-vessels no longer form exclusively the branchial organs; loose and large-celled tissue (fig. 126, a a) is superadded to the proper bloodvessels, which are far less in relative size than those in the former variety of branchiae; into the cells of this tissue the fluid of the visceral cavity insinuates itself, its course being marked by a slow motion. There exists, however, another point of structural difference between the branchial organs of this group and those of the former, viz., that wherever the fluid of the peritoneal cavity is admitted into the interior of the branchial organs, the latter are invariably supplied more or less profusely with vibratile cilia.

Fig. 125. Arenicola piseatorum.
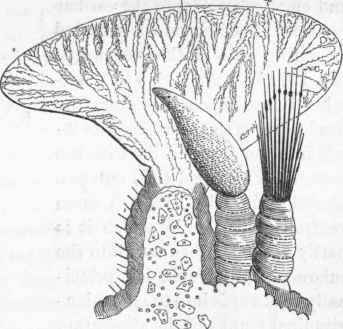
Fig. 126.
(667). In the branchial structure of worms thus constituted, the branchial appendages are found, instead of being composed of naked vessels, to present the appearance of round or laminated organs, into which the fluid of the visceral cavity freely penetrates.
(668). The blood-system is more concentrated in Arenicola than in any known Annelid. A large dorsal trunk (fig. 127, a; fig. 128, i), at the anterior three-fourths of the body receiving exclusively the efferent vessels of the branchiae, proceeds forwards from the tail and empties itself into the cardiac cavities, of which one is situated on either side of the oesophagus (fig. 127, b b'; fig. 128, b b.) Another vessel, proceeding from the head towards the heart, empties itself into the same cavity with the former. The blood then enters a second cavity (fig. 127, c' c'), more ventrally situated, by which it is partly propelled forwards into the suboesophageal trunk, but principally backwards into the great longitudinal trunks of the alimentary canal. The blood returning from the intestinal system of vessels reaches the dorsal intestinal (g) (lying in the median line underneath the dorsal trunk), from which the current diverges laterally at right angles into the branchiae (ff.) This conformation differs from that prevalent in all other Dorsibranchiate Annelidans, in which the great ventral trunk is the source of the branchial arteries. But the typical plan of the circulation is observed in the system of Arenicola at the posterior half of the branchial division of the body, where the afferent vessels of the branchiae emanate from the ventral trunk. It may be necessary to explain that the motion of the blood in that part of the circulating system which is anterior to the heart is the reverse of that posterior to this centre. The ventral oesophageal carries the blood forwards, and the dorsal backwards towards the heart.
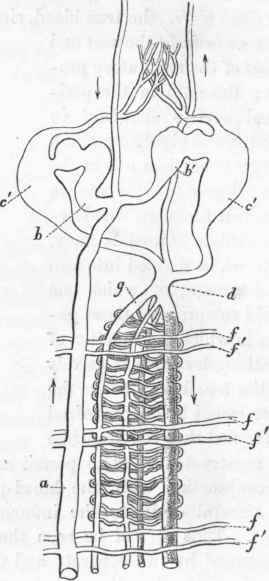
Fig. 127. Plan of the circulation in Arenicola. (After Dr. Williams).
(669). The independent contractile (ergo circulating) power of each individual vessel may be very completely proved by an examination of the branchiae of a living Arenicola. A single ramuscule in the branchial tuft may contract and empty itself while the surrounding branches are expanding diastolically. There is no synchronism in the circulatory movements of these vessels. Both the afferent and efferent vessels of the branchiae are long and tortuous, but discover no cardiac ampullae in any part of their course. In Arenicola the peritoneal chamber is filled with a highly corpusculated fluid, the basis of which consists of sea-water, and the presence and movements of which are indispensable to the circulation of the blood-proper. By this remarkable mass of fluid, the slender tortuous vessels are shielded from injurious pressure.
Continue to:
My Books

