Creatinin, The Measure Of Endogenous Protein. Metabolism
Description
This section is from the book "Modern Theories Of Diet And Their Bearing Upon Practical Dietetics", by Alexander Bryce. Also available from Amazon: Modern Theories of Diet and Their Bearing Upon Practical Dietetics.
Creatinin, The Measure Of Endogenous Protein. Metabolism
Despite this latter statement, creatinin may be taken as the measure of the antecedent tissue - or endogenous metabolism, and urea of the exogenous metabolism. Much experimental work must yet be undertaken to determine accurately the precise qualitative excretions of endogenous metabolism, and it is possible this exact estimation may never be completed, as it is so difficult to separate the two kinds of metabolism. If it were possible to make this calculation, then the answer to the question as to the necessity for this exogenous protein metabolism would be imminent. It is quite clear that enough protein to maintain endogenous protein metabolism is indispensable.
It has been assumed that the entire katabolism of protein takes place in the same tissues, i.e., chiefly the muscles, by oxidation in a similar manner to the fats and carbohydrates. But this is erroneous, as no oxidation is necessary for the decomposition of proteins, hydrolysis being sufficient. For this reason it is unnecessary to assume that all the kata-bolic processes take place in the muscles. Urea, the chief nitrogenous substance excreted in exogenous metabolism, is only found in extremely small quantities in muscle. On the other hand, creatin, the chief extractive of muscle, cannot be converted into urea either there or in the liver, but is eliminated as creatinin.
It is proper to add that the formation of creatinin is still a subject of great controversy, but, as will be more fully indicated in a later chapter, in all possibility the liver is responsible for its production. Creatin injected into the blood stream does not contribute to the formation of urea, but is excreted unchanged. Nor is there the slightest proof that the creatinin of the urine is derived from the muscular creatin. On the other hand, it is a fact of some significance that in diseases of the liver creatinin is excreted in diminished amount. It is conjectured that certain products of protein katabolism are carried to the liver, there transformed into creatinin, which is transported to the muscles as creatin. When the muscular store is complete the excess of creatinin is excreted by the kidneys. With a mixed diet from .8 grams to 1.2 grams of creatinin are excreted daily; with a vegetable diet, somewhat less, .6 to .86 grams.
It is perfectly evident from what has been said that by far the greater portion of the urea appearing in the urine is not formed in the muscles. We know that outside the body urea very quickly decomposes into carbonic acid and ammonia, to both of which substances it is very closely related. It is indeed the diamide of carbonic acid, as will be obvious from a study of its chemical composition.
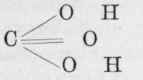
is carbonic acid. Replace one of the hydroxyl groups by an amide group, and we get which is carbamic acid, and by a repetition of the process we get which is carbamide or urea. On the other hand, ammonium carbonate is easily converted into ammonium carbamate by the loss of a molecule of water, and by the loss of another molecule of water becomes carbamide or urea.
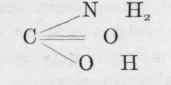
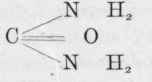
Now we have already seen that in protein digestion a certain amount of ammonia is formed, and we know that ammonia is produced in the organism from amino-acids. We also know from feeding experiments on rabbits and dogs that the body possesses the power of creating urea from ammonia and most of its salts, especially the carbonate, citrate, formate, and lactate, although the chloride, phosphate, and sulphate of ammonium are exceptions to this rule.
That the liver is the seat of this urea formation is considered a certainty from the following facts. In the first place urea ceases to be excreted in the urine and is replaced by ammonia when the liver is removed from frogs. A similar operation cannot be parformed in mammals because of its fatality, but on establishing Eck's fistula in the dog, whereby the blood of the portal vein is divided from the liver and passed directly into the vena cava, the amount of urea diminishes from about 90 to 77 per cent., and its place is taken by ammonia. In certain diseases of the liver, e.g., acute yellow atrophy, the urea practically disappears from the urine, whilst ammonia and the amino-acids, leucin and tyrosin, are present. Now we know that these originate in the intestine and on absorption pass through the liver, where they are usually converted into urea. Finally the blood in the portal vein is known to contain a larger quantity - nearly three times the amount - of ammonia than that in the hepatic vein, and by injecting defibrinated blood containing ammonium carbonate Schroder was able to obtain urea in abundance from the hepatic vein, no other organ in the body giving a similar result. It is therefore clear that exogenous nitrogen may become converted into urea without going near the muscles.
We also know that there is a limit to the growth of muscle in man, as only during the period of youth and convalescence, or to a certain extent during a holiday or severe physical exertion, is there any addition to the muscular system. A large quantity of food protein is not therefore required for building new tissues, because after attaining full growth the daily wear and tear of the tissues is small.
Yet we have seen that practically all the protein consumed is digested and absorbed, and if it be not used for building purposes, for what other purpose can it be destined? Now, it has always been assumed that the tissues use protein in preference to fats and carbohydrates, because the bioplasm has a large nitrogen content. If this be the case, then it follows that the more protein we swallow the better, a conclusion to which even high-protein advocates would not give their assent.
Continue to:
My Books

