Period Of Occurrence And Importance Of The Psychic. Part 2
Description
This section is from the book "The A. B. - Z. Of Our Own Nutrition", by Horace Fletcher. Also available from Amazon: The A. B.-Z. Of Our Own Nutrition.
Period Of Occurrence And Importance Of The Psychic. Part 2
We have now begun the analytical examination of the variations of our secretory curve. But owing to the importance of the matter we did not confine ourselves to conclusions which might be drawn from earlier investigations. We turned to new forms of experiment for further proof.
Thus we divided the ordinary ration of flesh given to our dogs - 400 grams - into four equal parts, which were administered at intervals of an hour and a half. (Experiments by Privat docent Kotljar and Dr. Lobassorff.) Each time after the dog received its 100 grams of flesh we were able to detect a rise both in the quantity and in the digestive power of the juice. The following table shows the figures in question:
Half-hour periods. | Quantity of juice. | Digestive power. | Remarks. |
1st | 3.1 C.C. | 5.13 mm. | 100 grms. flesh given. |
2nd | 5.0 " | 4.63 " | |
3rd | 4.7 " | 4.50 " | |
4th | 5.4 " | 4.88 " | 100 grms. " " |
5th | 5.5 " | 338 " | |
6th | 4.7 " | 2.75 " | |
7 th | 6.0 " | 3.75 " | 100 grms. " " |
8th | 5.4 " | 2.50 " | |
9th | 5.9 " | 2.50 " | |
10th | 5.4 " | 3.88 " | 100 grms. " " |
11th | 5.3 " | 3.0 " | |
12th | 4.2 " | 2.5 " |
In the curve which follows, only the variations of digestive power are represented.
It is clear that the increase, both of digestive power and of juice volume, is connected with the act of taking in food.
It appeared of interest definitely to determine the volume and properties of the secretion called forth by the act of eating in the dog with the isolated stomach. We endeavoured, therefore, at the beginning, to imitate the conditions of sham feeding as they occurred in the case of the dog with divided oesophagus. In addition to the fistular orifice leading into the isolated miniature stomach, another was opened into the main portion of the organ. If we now fed the dog in the ordinary way with small pieces of flesh, these were received back again at the orifice of the latter fistula, covered with saliva. Precisely as in sham feeding, after five minutes the juice began to flow simultaneously, from both the large and small stomachs. The secretion ran a corresponding course in the two cavities and ceased at the same length of time in both after the administration of food was stopped. Here is an instance taken from such an experiment performed by Dr. Lobassoff.
In five minutes the dog had eaten eighty pieces of flesh (weighing 172 grams), all of which soon afterwards dropped out at the fistula. The secretion began in both stomachs after the lapse of seven minutes from the commencement of the feeding, and proceeded as follows:
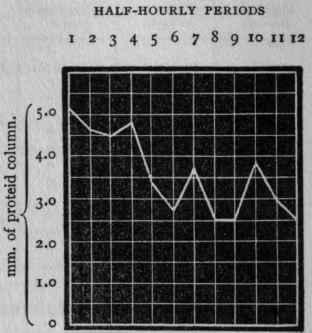
Figure 1. - Curve of digestive power constructed from the foregoing table.
Hour. | Miniature stomach. | Main stomach. | ||
Quantity of juice. | Digestive power. | Quantity of juice. | Digestive power. | |
I | 7.7 CC. | 83.2 CC. | 5.35 mm. | |
2 | 4.5 " | 6.25 mm. | 58.I " | In consequence of a mixture with bile (10-15 c.c.) the digestive power was greatly reduced. |
2 1/2 | 0.6 " | 8.5 " | ||
The secretion from both cavities also came to an end at the same time.
This experiment proves to us, first, that the main and miniature stomachs work in perfectly parallel manner with each other. The beginning, the end, and the intermediate variations of the secretion correspond in both cases. Secondly, the digestive power of the secretion coincides in both, and is the same which was observed in the so-called sham feeding. It has here remained at the same height till the cessation of the secretion, without falling to the lower value which we observed from the beginning of the second hour onwards, after normal flesh feeding.
This was also confirmed later, when we performed an cesophagotomy on the dog, and carried out sham feeding in typical form. Here follows one of these experiments taken from Dr. Lobassoff's article.
The first drop of juice appeared from both cavities during the sixth minute after commencing the feeding, which was kept up for half an hour. The further course of the secretion was as follows:
Hour. | Miniature stomach. | Main stomach. | ||
Quantity of juice. | Digestive power. | Quantity of juice. | Digestive power. | |
1st | 7.6 C.C. | 5.88 mm. | 68.25 C.C. | 5.5 mm. |
2nd | 47 " | 5.75 " | 41.5 " | 5.5 " |
3rd | 1.1 " | 5.5 " | 14.0 " | 5.38 " |
13.5 (total) | 5.75 (mean, | 123.75 (total) | 5.5 (mean) | |
The secretion came to an end in both stomachs at the same time.
The above is represented in curves in Figs. 2 and 3, the scale on which that for the main stomach is drawn being ten times less than that for the small. As you see, the progress of secretion is identical in both.
The existence of a fistula leading into the large stomach affords us also the possibility of performing an experiment upon our dog which is exactly the converse of the sham feeding experiment, and which constitutes a real experi-mentum cruets. While in sham feeding, we had only, so to speak, the beginning of digestion before us, we are now able in our cross experiment to start at the continuation of this beginning. For this purpose it is only necessary to bring the food into the stomach through the fistula, without attracting the dog's attention. Since in this experiment it is above all necessary not to excite the dog's appetite, it is best to carry out the procedure on the sleeping animal. I may add at once, however, that the same result can be obtained on the waking animal, only the process must be performed unnoticed, and the animal's attention must be diverted from thoughts of food.
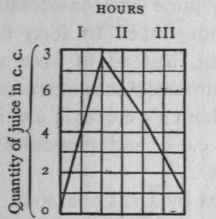
Figure 2. - Curve of secretion from the miniature stomach.
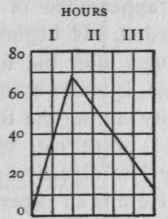
Figure 3. - The same from the main stomach reduced ten times.
The results of this experiment are striking, and do not in any way resemble the secretion after normal feeding. Some kinds of food, for instance bread and coagulated white of the hen's egg, when directly introduced into the stomach, do not yield a single drop of juice during the first hour or more afterwards. This holds good both for the small and large stomachs. When a glass rod is introduced into the food contained in the organ it remains dry. Flesh, if introduced at this stage, is able to excite a secretion, but the appearance of the juice is considerably retarded. It begins from fifteen to forty-five minutes after the feeding, instead of from six to ten, is under normal circumstances extremely scanty during the first hour (3 c.c. to 5 c.c. instead of 12 c.c. to 15 c.c), and possesses a very low digestive power.
Here is an experiment by Dr. Lobassoff:
400 grms. of flesh were brought into the stomach.
Hour. | Quantity of juice. | Digestive power |
1st . . | . 3.7 C.C. . . . | 2.0 mm. |
2nd . . | . 10.6 "... | . 1.63 " |
3rd . . | . 9.2 "... | . 1.5 " |
4th . . | . 7.0 " ... | . 1.88 " |
5th . . | . 5.6 " ... | . 2.25 " |
6th . . | . 6.6 "... | . 2.63 " |
7th . . | . 7-5 ". ... | . 1.88 " |
8th . . | . 5.3 " ... | . 2.0 " |
9th . . | . 3.0 " ... | . 5.0 " |
10th . . | . 0.2 "... | ____ " |
The secretion began twenty-five minutes after introducing the food. I now ask you to compare the following tables:
Hour. | Fed with 200 grins. of flesh (Chigin). | Flesh (150 grms.) brought into stomach (Lobassoff). | Sham feeding (Lobassoff). | Total quantity of juice in two experiments. | |||
Quantity of juice. | Digestive power. | Quantity of juice. | Digestive power. | Quantity of juice. | Digestive power. | ||
1st | 12.4 CC. | 543 mm | 5.0 CC. | 2.5 mm. | 77 c.c. | 6.4 mm. | 12.7 CC. |
2nd | 13.5 " | 3.63 " | 7.8 " | 2.75 " | 4.5" | 5.3 " | I2.3 " |
3rd | 7.5 " | 3.5 " | 6.4 " | 3.75 " | 0.6 " | 5.75" | 7.0 " |
4th | 4.2 " | 3.12 " | 5.0 " | 3.75 " | - " | -" | 5.0 " |
The progress of juice secretion in the above is also represented in the following curves:
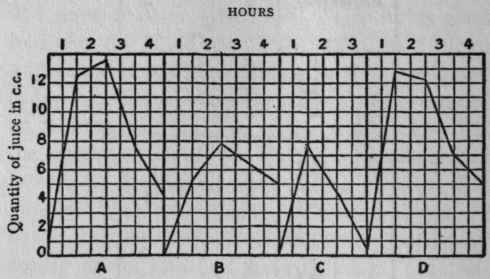
Figures 4-7. - A. Ordinary curve of gastric secretion (200 grms. flesh). B. Curve from direct introduction of food (150 grms. flesh). C. Sham feeding with same. D. Summation of B and C.
As you see, the curve which represents the results of the direct introduction of flesh ascends much more slowly and does not attain anything like the height of that caused by normal feeding with the same food. But if the quantities obtained by direct introduction of the flesh be added to those of sham feeding, the resulting curve is almost identical with the normal.
In like manner the digestive power of the secretion in the foregoing experiments can be dealt with, and with the same result. It is a good instance of how a secretion curve can be synthetically constructed from its constituent factors.
Continue to:
My Books

