Chapter XXVIII. General Physiology Of The Nervous System
Description
This section is from the book "A Manual Of Physiology", by Gerald F. Yeo. Also available from Amazon: Manual Of Physiology.
Chapter XXVIII. General Physiology Of The Nervous System
Anatomical Sketch
The nervous system includes the various mechanisms by which the distant parts of the body are kept in functional relationship with one another. By it the condition of the surroundings and the various parts of the body are communicated to a central department (cerebro-spinal axis) which in turn regulates and controls the activities of the various organs.
It is made up of two varieties of tissue, both of which possess special vital properties. The one, nerve fibres, composed of thread-like strands of protoplasm, connects the elements of the other, nerve corpuscles, which form the peripheral or central terminals of the fibres. Nerve fibres are simply special conducting agents, having at one extremity a special terminal, or nerve cell, for sending impulses, and at the other end a nerve cell for receiving the impulses. These terminal organs, between which the nerve fibres pass, are the agents which determine the direction in which the impulse is to travel along the nerve. The sending organ may be at the peripheral end of the nerve, and the receiver in the nerve centres, as in the case of an ordinary cutaneous nerve, which carries impulses from the skin to the brain; or the sending organ may be at the centre, and the receiving organ at the periphery, as in the case of the nerves conveying impulses from the brain to the muscles.
The former kind of nerves are called afferent, carrying centripetal impulses, and the latter efferent, carrying centrifugal impulses. Nerves are capable of carrying impulses in either direction, as has been proved by cutting the afferent lingual and the efferent hypoglossal nerves, and causing the proximal end of the former to unite with the distal end of the latter, which is distributed to the muscles of the tongue. When the union has taken place, a stimulus applied to the upper part which was normally afferent, or sensory, carries motor impulses to the muscles, i. e., acts as an efferent nerve.
Protoplasm, though not formed into fibres, can conduct impulses, as is seen in the transmission of an impulse in textures and animals which seem to have no special conducting elements or nerve fibres. Thus, in the hydra all the cells act as nerves, and in the higher animals an impulse, producing a wave of contraction, can pass from one muscle cell to the other directly, as is seen in the ureter, or in the heart of cold-blooded animals.
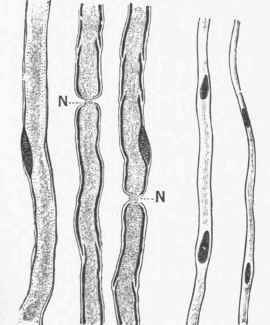
Fig. 200. Highly-magnified view of three medul-lated and two non-medullated nerve fibres of frog, stained with osmic acid, which makes the medullary sheath black.
N. - Nodes of Ranvier - where the axis cylinder can be seen to pass the gap in the medullary sheath.
Fig. 201. Transverse section of nerve fibres, showing the axis cylinders cut across, and looking like dots surrounded by a clear zone, which is the medullary sheath. Neuroglia separates the fibres into bundles.
The only essential part of a nervous conductor is a delicate protoplasmic fibril. Single, thin, thread-like fibrils are found carrying impulses in the nerve centres. In the nerves distributed about the body, one does not meet these single protoplasmic threads (except where the fibrils are interwoven to form terminal networks, as seen in the cornea), but the fibrils are clustered together in large bundles, so as to make one nerve fibre. In the peripheral nerves this bundle of protoplasmic fibrils is covered, 42 and is called the axis cylinder of the nerve fibre. In some nerve fibres there is but one very thin transparent covering, termed the primitive sheath, while in others there is a thick layer of doubly-refracting fluid inside the primitive sheath, in immediate contact with the fibrils of the axis cylinder. This is called the medullary sheath, or white substance of Schwann, because its peculiar refractive properties make it look white when viewed in a direct light. As the nerves have or have not this medullary sheath, they have been termed "white" or "gray." The former are by far the most plentiful, since they make up the greater part of the ordinary nerves, while the gray fibres only predominate in the sympathetic nerve and its ramifications, and parts of the special sense organs.
An ordinary nerve, then, is made up of a large number of fibres, held together by connective tissue, each fibre containing a vast number of fibrils within its sheath.
Functional Classification
Nerve fibres may be classified, according to their function, in the following way: -
I. Afferent Nerves, which bear impulses from the surface to the nervous centres. These may be further divided into: -
(a) Sensory nerves, when the impulse they convey gives rise to a "perception." The perceptions may be the special sensations which are transmitted from the organs of special sense, or those of general sensation, giving rise to pleasure or pain.
(6) Excito-reflex nerves communicate impulses to central nerve elements, and give rise to some action, without exciting mental perception. Such nerves regulate the viscera. According to the result of the excitation arising from their impulse, they are termed excito-motor, excito-secretory, and excito-inhibitory, etc.
(c) Mixed nerves act as sensory and reflex nerves; these are the most numerous, the sensory or reflex action depending upon the condition of the nerve centres.
II. Efferent Nerves, which carry impulses from the centres to the various organs throughout the body. According to the effect produced by their excitation, they are termed: -
(a) Motor, conveying impulses to muscles and exciting them to contract.
(/B) Secretory, the stimulation of which calls forth the activity of a gland.
(r) Inhibitory, when they check or prevent some activity by the impulses which they carry.
(d) Vasomotor nerves, which regulate the contraction of the muscular coat of the blood vessels.
(e) Trophic, thermic, electric nerves are also to be named, the two former being of doubtful existence, and the latter being only found in those animals which are capable of emitting electric discharges, such as electric fishes.
III. Intercentral Nerves act as bonds of union between the several ganglion cells of the nervous centres, which are connected, in a most elaborate manner, one with the other. The terminals of these fibres are possibly both receiving and directing agents, and the delicate strands of protoplasm communicating between them probably convey impulses in different directions, but of this we can have no definite knowledge, although such a supposition would aid us in forming a mental picture of the manner in which the wonderfully complete intercentral communications are accomplished.
Mode Of Investigation
In order to understand the functions of the different nerves a knowledge of their central connections and their peripheral distribution is necessary. But anatomical research, unaided by experimental inquiry, does not suffice to determine their function.
The procedure adopted in testing the function of a nerve is the following: The nerve is exposed and cut, and it is observed whether there be any loss of sensation or muscular paralysis in the part to which it passes. The end connected with the centres is spoken of as the central or proximal end, and that leading to the distribution of the nerve is called the peripheral or distal end. Each of these cut ends is then stimulated, and the results are observed. If the nerve be purely motor, stimulation of the proximal end will yield no result, but when the distal end is irritated, movements follow. If, on the other hand, it be a sensory nerve, stimulation of the distal end gives no result, and that of the proximal end produces signs of pain.
Chemistry Of Nerve Fibres
The axis cylinder of nerves is probably composed, as already mentioned, of protoplasm; further than that nothing is known of its chemical properties. The medullary sheath yields certain substances which are related to the fats, and can be extracted with ether and chloroform. Among these is the peculiar compound nitrogenous fat, lecithin, containing phosphorus, also cholesterin, cerebin, and kreatin.
Electric Properties Of Nerves
Like muscle, nerves may be regarded as having a state of rest and a state of activity, but the two states are not obvious in the same striking way as they are in muscle, nor do we know much of the physical properties of nerve. While at rest, however, it shows electric phenomena similar to those which have already been described as belonging to muscle tissue. These electrical currents are contemporaneous with the life of the nerve, and they undergo the same variation as occurs in muscle when the nerve passes into the active state; that is, when it transmits an impulse.
The so-called natural current of nerve is practically the same as that of muscle, passing in the nerve to the central part from the cut extremities of the fibre; that is to say, the current passes through the galvanometer from the electrode leading from the middle of the nerve, to that applied to the extremity. The electromotive force of a small nerve is much less than that of a muscle. In a frog's sciatic it has been estimated to be 0.02 of a Daniell cell. The natural current of the frog's nerve is said to increase in intensity in proportion to the increase in temperature up to about 200 C, after which it decreases.
Experiments on nerve currents must be carried on with all the precautions mentioned in speaking of muscle currents, and with the non-polarizable electrodes there figured (page 448).
Continue to:
My Books

