Inhibition Of Reflex Action
Description
This section is from the book "A Manual Of Physiology", by Gerald F. Yeo. Also available from Amazon: Manual Of Physiology.
Inhibition Of Reflex Action
The great majority of reflex actions may be prevented or controlled by the will, and the basal ganglia and medulla habitually exert a checking or inhibitory influence on the reflex actions of the spinal cord. It is in this way that we account for the facts that a living frog when stimulated does not respond with the ordinary reflex movements, and that a human being, when asleep, shows reflex action in response to a slight stimulus that would be quite ineffectual were he awake. For some little time after pithing a frog, constant or regular results are seldom met with, because the section of the upper part of the spinal cord acts as a stimulus to those channels which habitually bear impulses from the brain, and, by exciting them, has inhibitory effect. Further, artificial stimulation of the corpora quadrigemina and medulla have the effect of checking the reflex action of the cord.
If, while the cord is employed in reflex action, in response to gentle cutaneous stimulation, the central end of a large sensory nerve trunk be stimulated, the reflex action ceases. In short, it may be accepted that strong impulses arriving at the cord from any direction have the effect of inhibiting the action of its reflecting cells.
The theory of reflex action lies at the bottom of all nervous activities, and it is therefore useful to attempt to work out the details of the mechanisms by means of which it is carried on. A simple plan of the channels traversed by the impulses is given in the diagram (Fig.' 249), in which the arrow heads show the direction of the afferent impulse passing along the posterior root to reach a cell in the posterior gray column, thence it passes to a cell in the anterior column, to reach the efferent fibre, and through the anterior motor root of the nerve on its way to the muscle. It has been suggested that the impulse meets with considerable resistance in passing through the protoplasm of the cells, and that owing to this resistance, the effect of a slight stimulus remains localized, while more powerful impulses can overcome the resistance, and spread to a greater number of cells. Thus, the regular radiation in the cord would be simply dependent on the inability of the impulses to affect cells other than those in their immediate neighborhood. Following out this view, it has been suggested that the resistance is increased by impulses arriving at the cells from a different direction, and the inhibitory action of the higher centres, or peripheral excitation of another part, impedes the spreading of the impulses.
But this theory of resistance to and interference with the transmission of impulses in the nerve cells hardly explains all the phenomena observed in the reflex action of the spinal cord and the various modifications it can undergo.
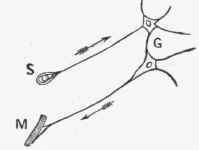
Fig. 249.
S. Sensory receiving organ with attached afferent nerve fibre.
G. Central organs - ganglion cells.
M. Peripheral organ and efferent nerve.
The reflex convulsions that occur in poisoning with strychnine, or as the result of some constant but slight stimulation, may be explained as follows: -
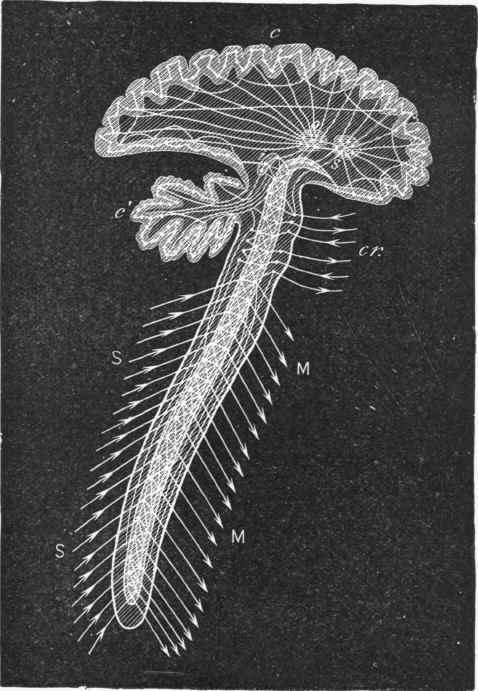
Fig. 250. Diagram of the paths taken by the impulses in the brain and cord. MM, motor channels; SS, sensory channels; cr, cranial nerves.
Besides the resistant protoplasmic fibrils in the gray part of the cord, there exist medullated fibres in the root zones - short cuts, as it were - by which impulses travel from one part of the cord to another. If we suppose the ordinary reflex traffic of the cord cells to be carried on without the assistance of these direct lines of communication, we must assume that there is some special means of shutting these fibres out of the working of the reflex machine. Such special mechanisms in all probability exist, and are in relationship with or under the command of the inhibitory cells of the higher centres. We may then suppose that strychnine removes the power of these inhibitory agents, and the impulses finding the direct ways open, take these routes, and are simultaneously and irregularly diffused throughout all the cell territories (independent of the ordinary paths they have been educated to follow), and thus convulsive movements are excited in many parts of the body.
In like manner the unremitting activity necessary to keep in check the impulses arriving from a constant source of stimulation (such as intestinal worms), eventually fatigues the active elements in this inhibitory mechanism, and then - often suddenly - the force of the accumulated irritation rushes along the direct channels to all parts of the cord, and simultaneously exciting them, brings many discordant muscles into spasmodic action.
The reflexion of an impulse from a sensory nerve, through the cells of the spinal cord to a motor nerve, occupies a measurable length of time, which has been estimated at about 1/10 of a second. The time required for the performance of a reflex act varies considerably in the same individual under different conditions; of these, high temperature and intense stimulation shorten the time, and fatigue or cold lengthen it.
Continue to:
My Books

