Lipidic System
Description
This section is from the book "Research In Physiopathology As Basis Of Guided Chemotherapy With Special Application To Cancer", by Emanuel Revici. Also available from amazon: Research In Physiopathology
Lipidic System
The relative insolubility of the lipids in water and their solubility in neutral solvents has permitted us to separate these substances as a group from the other constituents of organisms. For more than just didactic purposes, we consider lipids to constitute a separate system in the organism. The part played by lipids in the organization and the functioning of various entities supports this concept. For example, when a lipoid is introduced into the organism, it will be selectively dissolved in, circulated through, retained by and metabolized as part of the lipidic system. Overtone's "Index of Repartition" of anesthetics in the organism can be seen to be a direct corollary of the existence of such a system although the anesthetic agent can be a lipoid or a nonpolar substance.
A great degree of independence of this system is morphologically evident as in adipous cells, when fats circulate as chylomicrons or when they form oriented layers. We have seen above how the orientation of lipoids at the surface of water results from the relationship between the solubilities of the two constituent groups, polar and nonpolar. Along with their insolubility in water, the orientation of lipoids has allowed them to play a very important role in biology.
The very existence of biological entities appears to depend upon the ability of lipids to build up boundary formations separating and thus assuring the individuality of biological entities.
Through peculiar, reciprocally opposed orientations, two or more layers of lipids can form a membrane with two polar faces which has the ability to separate two aqueous media. In its simplest form, such a membrane appears in mitochondria. (Note 3) Similar boundary formations identify nuclei and cells and appear in higher entities, as in the membranes and intercellular cements of lymphatic and blood vessel endothelia. It is this peculiar orientation which allows lipids to establish the necessary boundary formations resulting in complex hierarchic organisms. The existence of biological entities, at least from the chromosome level up (and probably even below that level), can be seen to result direcdy from the intervention of lipids as a separate system, particularly in the formation of the dipolar lipidic boundaries.
However, boundary formations which separate the biological entities would not have been efficient if they did not fulfill another capital role: that of allowing selective passage of metabolites. A totally impermeable membrane would isolate the respective entities and result in their death. On the other hand, a totally permeable membrane would have no usefulness. The boundary formation has to act selectively, permitting the passage of some, but not all, substances. But even this does not seem to be sufficient to insure an efficient boundary. Most important, such a membrane must be able to alter its permeability, quantitatively and qualitatively, according to variations in circumstances. Such capacity for altering permeability can be related to the presence of the two groups of lipids, fatty acids and sterols, with their antagonistic properties relating to permeability.
The fatty acids appear to induce permeability in the membrane they form, especially permeability for anions. The perpendicular position to the surface of water assumed by the nonpolar aliphatic groups when the fatty acids form this boundary membrane appears to be favorable for the passage of a substance through the membrane. The fatty acid molecules thus can be separated, permitting other molecules to pass between them; that is, to pass through the membrane formed by the fatty acids. The negative electrical character of the polar groups of these fatty acids explains why they represent a kind of barrier to the free passage of cations. These cations are attracted and retained by the acid polar group. This would explain the manifest changes in permeability under the influence of calcium ion. The removal of calcium from cellular membranes, through treatment with oxalates, increases permeability, while treatment with calcium salts reduces permeability. The bivalent calcium ion, when it binds the polar groups of two adjacent fatty acid molecules in the membrane, prevents the passage of other molecules between these parts of the membrane, a fact which explains the manifest decrease in permeability induced by this cation.
The other group of lipids, the sterols, have an effect on permeability opposite to that of fatty acids. This can be related in part to the bond which these sterols make with the fatty acids. Consequently, they block any passage through the part of the membrane formed by fatty acids. The impermeability is due, to some extent, to a peculiarity of the layers formed by these sterols themselves. The polycyclic molecules of sterols do not take the same perpendicular position toward the surface of water as fatty acids do. (27) (Fig. 63) Since sterol molecules assume a position almost parallel to this surface, the layer which they form exhibits no permeability properties. It even opposes any passage through it. It seems that fatty acids and sterols make separate "spots" in the cellular membranes so that, through their quantitative relationships, they confer different degrees of permeability to different regions of the membrane. The changes in permeability which result from the antagonistic intervention of the two groups of lipids seem to play an important role in normal and abnormal physiology.
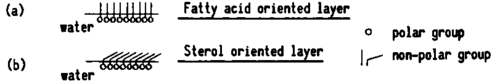
Fig. 63. Schematic aspect of oriented interface layers. The perpendicular position to the surface of water of fatty acid molecules (a) favors the passage of other molecules through the separating membranes they form. Oppositely, the almost parallel position to the surface of water of the polycyclic molecules of sterols (b) prevents the passage of other molecules through the membrane they form.
With fatty acids inducing permeability, and sterols opposing it, the fundamental character of their biological relationship can be recognized. It would seem that part of the function of sterols is to oppose the activity of fatty acids. Conceptually, sterols would appear, in this specific activity, to be "anti fatty" acid agents, with a capacity to control the activity of fatty acids rather than be active by themselves. Partly for this reason as well as for greater general understanding, it is necessary first to investigate fatty acid activity.
Continue to:
My Books

