Biological Problems In The Production Of Tumors And The Design Of Experiments
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Biological Problems In The Production Of Tumors And The Design Of Experiments
Tumors induced by viruses simulate naturally occurring spontaneous tumors in that they derive entirely from the hosts' own tissues. They are therefore devoid of divergent histocompatibuity antigens which sometimes arise through gene mutations when transplanted tumors are maintained for long periods, even in highly inbred strains of animals, and which may contribute to extraneous antitumor effects that are not observed with indicator tumors of the naturally occurring variety.
Two things are essential to the use of viral tumors in large-scale testing programs: (1) There must be an abundant and economical source of virus of known, or standardized, potency so that there will be no variation in the initiating dose in the production of large experimental lots of indicator tumors. (2) Populations of test animals must be available which are highly susceptible to the virus, and in which the variations among individual animals follow a fairly uniform and predictable distribution about their averages with respect to certain measurable biological properties, such as time elapsing between inoculation of virus and the appearance of a tumor, rate of growth of the tumor, and time-to-death caused by the tumor.
When tumor viruses are first isolated from their natural hosts, some difficulty is usually experienced in obtaining reproducible results on re-inoculating the virus into indicator hosts. The reasons for this are varied and are not completely understood, but in several established systems the particular combinations of virus-strain and test-animal population are sufficiently adapted to each other, and the interactions between host and virus in relation to dose are sufficiently stable to permit quantitatively controlled experimentation with chemotherapeutic agents. In the systems in which such stability exists, the data accumulated for demonstrating it can also be used as a basis for planning tests of therapeutic and prophylactic effects.
The information needed is the same as that required as background for the development of bioassay methods based upon various biological criteria of response. Thus, in any system it is necessary to know first the dose range within which the incidence of positive responses, i.e., tumors, among groups of inoculated animals decreases from 100 to 0 percent, as well as the dose which on an average just produces, or closely approaches, a 100 percent incidence. It is the relation of this latter dose (in terms of its dilution factor) to the undiluted virus suspension which determines the practicability of a particular virus-host system for the large-scale production of indicator tumors. An example is apparent from a consideration of text-figure 1, the right chart of which illustrates the usual sigmoid percentage-incidence curve obtained with serial dilutions of a virus. The curve represented was obtained with Rous sarcoma virus (2). In this experiment the highest dilution giving a 100 percent response was 10^-5, but in many experiments carried out over a long period it has
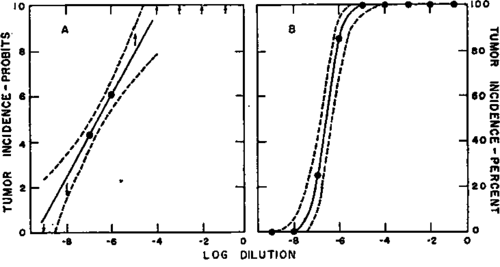
Text-figure 1. Responses of homogenous groups of chickens to the Rous sarcoma virus-see text. (Reproduced from J Nat. Cancer Inst. 16: 856, 1956.) been observed to vary from about 10"6 to about 10^-4, due to fluctuations in test-animal susceptibility (S, 4). A safe dilution for the routine production of indicator tumors would be 10^-4, with a high probability of obtaining tumors in most animals inoculated. Since the log dilution values shown along the abscissa of text-figure 1 are in terms of gram equivalents, the zero value at the right represents the quantity of virus derived from 1 gm. of tumor tissue, suspended in 1 ml. of diluent. The constant volume used for inoculating the chickens was 0.2 ml. for each of the dilutions shown. Thus, if only the 10^-4 dilution were to be used for chicken inoculations, 1 ml. of the original suspension would provide 10 liters of the desired concentration, or enough for the inoculation of 50,000 test animals. This information is of importance, not because it would ever be desired to inoculate such a large number of animals at any one time, but because the figure represents the total number which eventually could be inoculated with the amount of virus derived from 1 gm. of source tumor tissue, because the virus remains stable upon freezing and storage at temperatures below - 50 C. (8). It therefore gives a clue to the magnitude of the problem that would be involved in producing adequate quantities of standardized seed virus for use by many laboratories over long periods. In the example cited, each ml. of the original virus suspension could be further divided into 10 subsamples of 0.1 ml. each, for sealing in separate glass ampules, and each ampule would provide enough virus for the inoculation of 5,000 animals; or the original suspension could be diluted 1:10 and 100 subsamples prepared from 1 ml., each being enough for the injection of 500 test animals. Since the processing of 100 gm. of tumor tissue in the preparation of virus suspension is not an undue burden for the average virus laboratory, it is not the preparation of virus itself but the parceling of virus into subsamples that represents the limiting factor in the production of seed virus in most research laboratories. If industrial facilities were available for parceling, identical subsamples of a common virus preparation could be made available in lots of tens of thousands with this particular virus. A given 100 ml. lot of virus suspension would therefore provide enough samples for the eventual inoculation of 5 million chickens for the induction of indicator tumors. The use of such standardized lots is of primary importance in work with the tumor viruses because major fluctuations occur in the yields of virus from different source lots of tumor tissue, and virus suspensions prepared at different times by the same techniques cannot be counted upon to be quantitatively comparable with respect to potency. However, the potency relationships between different lots can be readily established through bioassays.
The left chart of text-figure 1 shows in probits the percentage results given in the right chart. This illustrates the application of a simplified biometric procedure for estimating the dose that would be expected
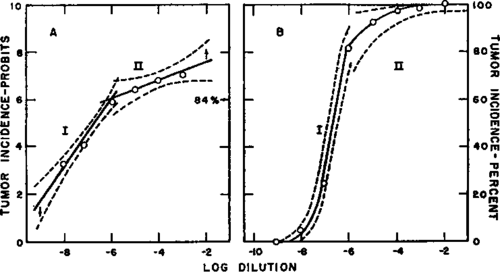
Text-figure 2. Responses of heterogenous groups of chickens to the Rous sarcoma virus-see text. (Reproduced from J Nat. Cancer Inst. 16: 859, 1956) to produce any given percentage response (2). Since the approach to 100 percent is asymptotic, the dose estimated to yield a 99 percent response (7.3 probits) may be considered as that which "just produces" a 100 percent response (5).
Most strains of chickens are not entirely homogeneous genetically (see 6-8), and some lots of test animals will contain individuals of aberrantly high resistance. The dose required to induce tumors in all the individuals, including the highly resistant ones, would therefore be unrealistically high. In this case the percentage results are truncated, as illustrated in text-figure 2, and the dilution to be used for routine production of indicator tumors is estimated by extrapolation of the initial, steep portion of the probit curve to the 99 percent (7.3 probit) level (see 9) rather than by taking the dilution actually required.to produce a 100 percent incidence. There will, of course, be some loss in experimental animals in this situation since all of them will not develop tumors with the particular dose employed.
Continue to:
My Books

