Constitution Of Tumor Viruses. Part 3
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Constitution Of Tumor Viruses. Part 3
According to either of these theories, a situation is envisaged in which the old idea that tumor viruses merely trigger off malignant change as a side effect during replication would, after all, be justified.
Plate 6
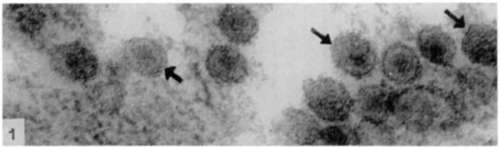
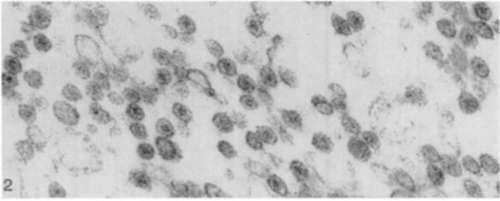
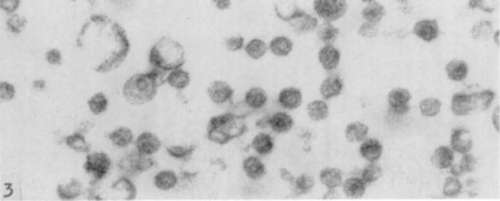
All the figures are electron micographs of sections cut through permanganate-fixed samples from the virus-containing zones of pellets prepared from fluoro-carbon-treated Rous virus suspensions.
Figure 1. Group of particles about 75 m/i in diameter showing electron dense nucleoid and less dense outer zone or viroplasm around it. The double nature of outer-limiting membrane is evident in particles most favorably orientated to plane of section (arrows). [Reprinted from the British Journal of Cancer; Epstein (13).] X 150,000
Figure 2. Survey picture of control sample which had been incubated for 2 hours at 37° C. in 30 percent alcohol. The spherical virus particles appear ragged and somewhat extracted; the outer-limiting membranes and dense nucleoids are intact. [Reprinted from the British Journal of Cancer; Epstein and Holt .(10).] X 70,000.
Figure 3. Survey picture of sample incubated as that shown in figure 2 except that the alcohol contained ribonuclease. The nucleoid has been removed from each particle, whereas the viroplasm and outer-limiting membranes have not been affected. Particles appear slightly swollen as if some central bracing structure has been removed. [Reprinted from the British Journal of Cancer; Epstein and Holt (16).] X 70,000.
Figure 4. Low power survey picture showing composition of virus-containing zones of pellets. Uniform particles about 75 mµ in diameter are almost the only formed structures present. [Reprinted from the British Journal of Cancer; Epstein (13) .1 X 14,000.

Discussion
Dr. Mellors (Hospital for Special Surgery, New York): I wish to compliment Dr. Epstein on his fine presentation. He has wisely extended his work from an RNA-virus such as Rous sarcoma virus to a DNA-virus such as adenovirus. In considering the contents of his submitted report, I was a little concerned about the purity of the virus-containing pellets that are obtained by the fluorocarbon and high-speed centrifugation method, wondering, for example, in the case of an RNA-virus pellet whether cytoplasmic particulates containing RNA might constitute a significant contaminant.
Dr. Epstein's work was done with such thoroughness that I was sure he had an adequate explanation for this, but the extension of the study into the DNA category with such clear-cut results puts all concern to rest.
I would like to ask him, however, if he has any idea as to why acridine orange stains and fluoresces in a certain color upon combination with DNA and in another color on combination with RNA. Is this just simply a matter of the concentration of the fluoro at a particular locus, a kind of metachromatic situation? Finally, you did indicate some problems in nonspecificity even in the staining of adenovirus. You overcame this problem effectively and properly but I would like to have a little clarification as to the nature of this problem.
Dr. Epstein: Well, I am no chemist, so I do not think I am going to be able to help you very much. With regard to the first question about the specificity of this staining reaction, the finding of acridine orange and its properties came as the culmination of an immense amount of work with various kinds of fluorochromes. The first indication of the specificity of the dye for nucleic acids was given in 1956, independently in my country by Dr. Armstrong, and in this country by Dr. von Bertalanffy working with Dr. Bickis. The way their two papers appeared almost simultaneously with identical results and the subsequent work which has been done with the technique make it quite clear that for some reason which may be known to the chemists but which is not known to me, this fluorescence response is absolutely specific for the two types of nucleic acid, if you confirm it with a nuclease digestion.
If you can abolish the fluorescence with a nuclease digestion, the specificity is sound. Why occasionally-and this is the second question-you should get a nonspecific fluorescence we do not know. We think it is some sort of technical thing, but it can be circumvented by insistence on a nuclease control. And if you do that, you seem to be all right.
Dr. Dmochowski (M. D. Anderson Hospital): I will first mention the question of acridine orange which Dr. Mellors has raised. I think, Dr. Epstein, when we speak of the specificity of staining with acridine orange we have to consider two types of specificity: (1) the specificity for DNA or RNA of the host, and (2) the specificity for DNA or RNA of the virus.
Dr. Niven and Dr. Armstrong in England and Dr. Tenenbauin in this country have pointed out that in viral infections you may get a certain degree of acridine orange staining which will be either for DNA or RNA, but this does not necessarily mean that it is the viral DNA. Whether it is of viral or host origin can be tested with comparative ease by pretreatment with chymotrypsin or pepsin followed by DNase.
Staining with acridine orange for DNA may be only host response, and in this case it can be removed by digestion with DNase. If it is due to the virus itself, it cannot be removed by DNase alone, but only after treatment with pepsin or chymotrypsin followed by DNase digestion.
I would like to ask Dr. Epstein about the sections of adenoviruses. How many times did you apply the fluorocarbon treatment to the adenovirus preparations before you obtained such nice sections? Are these sections of the whole pellet or of the so-called outer zone of the pellet, i.e., the upper zone of the sedimented pellet?
Dr. Epstein: Now, let us take those in order. The first one, I did not express any opinion at all as to whether the acridine orange was showing us that there was viral DNA or RNA-viral nucleic acid or host cell nucleic acid. All I said was that it is specific for nucleic acid.
The problem of what nucleic acid you are looking at is a completely different one and takes us into the field of virus isolation and purification.
The question of applying various other enzymes beforehand to test whether there is a viral or host cell nucleic acid applies only when you are looking at viruses inside a cell. It does not apply in the kind of material which we have examined because we have spun down purified virus particles, but whether there is, in addition, contaminating host cell material is another matter. If the material stained with acridine orange is checked and looked at in the electron microscope, morphological information can be obtained as to the degree of purification that has been achieved. This does not tell you whether other substances are present which cannot be seen with the electron microscope. But we think that, taken in conjunction with the electron microscope changes that are seen inside the virus when the digestion is carried out, we are justified in concluding that the material which we stained consists of virus particles.
You asked how many times we had treated the active virus with fluorocarbon, and the answer is three.
Another question was, Did this section represent the whole of the pellet? No, I described what we looked at as the virus-containing zone. There are other substances that are not removed by fluorocarbon and which come down in the pellet. However, if a Spinco centrifuge with an angle head is used, the various deposits separate out in different regions of the pellet, and an area made up of particles is seen.
That is a very extensive area and the uniformity which I showed you is not just an example of a good part; it is what is seen right through.
Continue to:
My Books

