Different Aspects Of Nucleoli In Cancer Cells
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Different Aspects Of Nucleoli In Cancer Cells
Figure 4. Nucleolus of Rous virus-infected cell grown in vitro. Note "moire" appearance of "nucleolonema" due to densification of some of its segments and accumulation of "pars amorpha" into small pools. X 15,000.
Figure 5. Pronounced condensation of nucleolar granules in human breast cancer- a frequent finding observed, also, in virus-induced tumor cells. Note "dense bodies" at (→). X 20,000
Figures 6 and 7. Other examples of nucleoli in human breast carcinoma. With exception of figure 7, other aspects are not usual in normal cells, yet they probably correspond to modified metabolism and cannot be considered as specific. X 11,000.




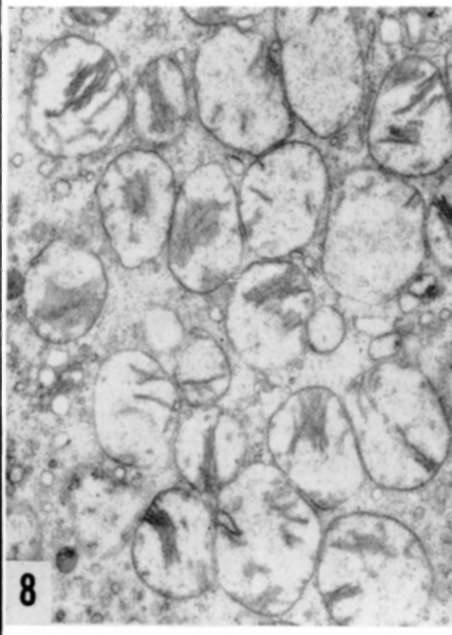



Plate 29
Figure 12. Portions of 2 adjacent cells from human breast carcinoma. Mitochondrial swelling is illustrated at right and presence of intracytoplasmic canaliculae with hypervillosity is seen at left. These two features are frequently encountered in these malignant cells. X 18,000.

Plate 30
Figures 13, 14, and 15. Different aspects of increase in ergastoplasm in Rous virus-infected cells. Accumulation of ribonucleoprotein granules in cytoplasmic vacuole (fig. 13), hyperplasia of lamellar (fig. 14), and granular (fig. 15) ergastoplasm in cytoplasm. X average 33,000.
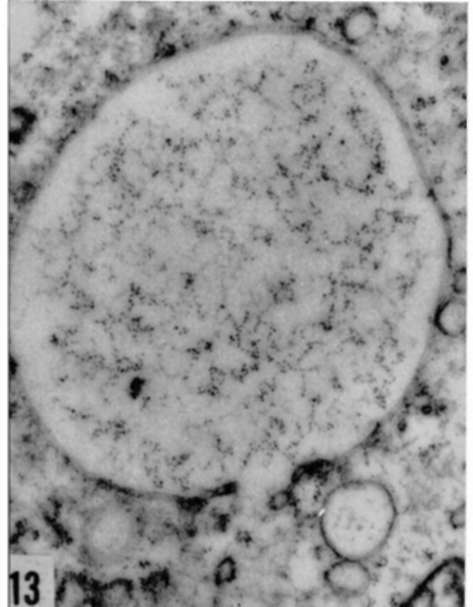
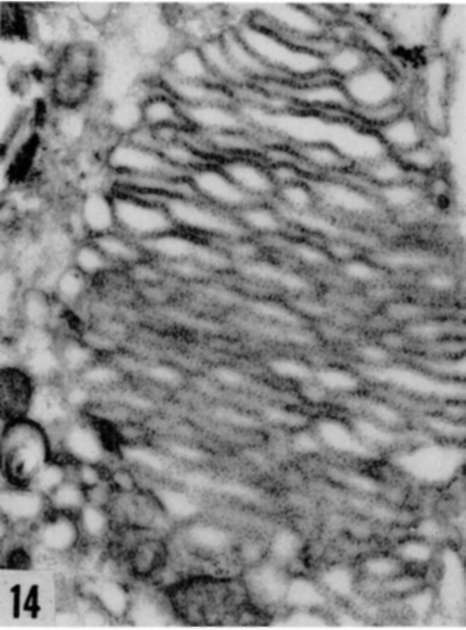
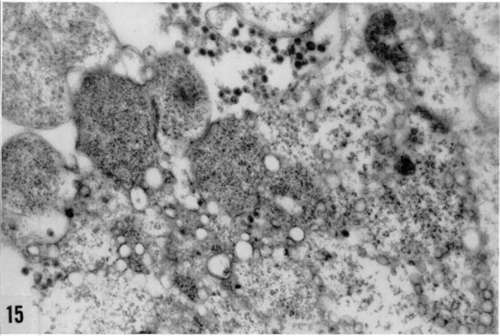
Plate 31
Figure 16. Remarkably hypertrophied cellular center in Rous sarcoma virus-infected cell observed in optical microscope. X 2,000.
Figure 17. Ultrastructure of this region to show that it corresponds only to packing of Golgi vacuoles (→). No virus particles are associated with such hypertrophied Golgi apparatus. X 18,000.
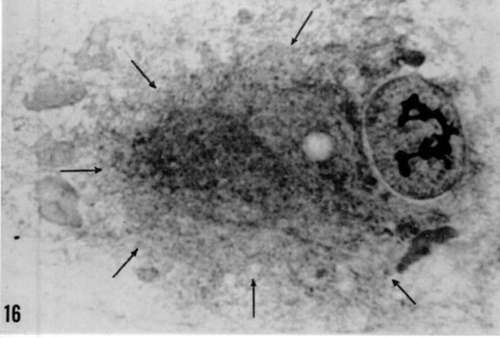

Plate 32
Diverse particulate formations to illustrate difficulties sometimes encountered in morphologic identification of viral particles. Only particles in figure 21 may be identified as such.
Figure 18. Typical "compound vacuole" in testis of guinea pig (courtesy of J. Andre). X 42,500.
Figure 19. Particle-containing vacuole in human breast cancer. Significance of structure is unknown. X 30,000.
Figure 20. Nuclear "pores" from very indented nucleus in Rous sarcoma cell. X 40,000.
Figure 21. Purified preparation of Shope papilloma virus particles. X 120,000.




Plate 33
Figure 22. Increase in cytoplasmic vesicles (endoplasmic reticulum) in Rous virus-infected cell. X 51,000.
Figure 23. Pinoeytosis vesicles and their relationship to surface membrane and to intracellular vacuole in Rous virus-infected cell. Virus particles have probably been engulfed through this process. X 40,000.
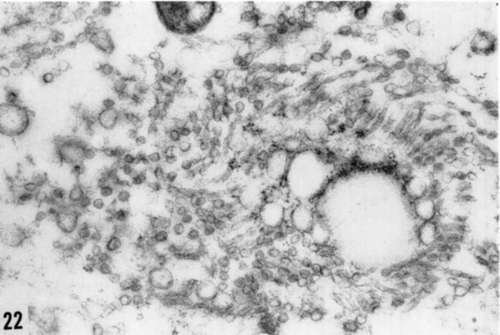

Plate 34
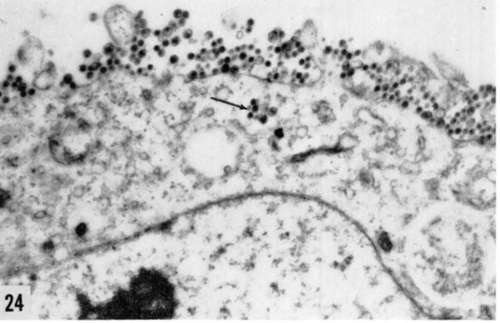
Figures 24, 25. and 20. Extracellular (fig. 24) and intracellular (figs. 25 and 26) types of virus particles found in cells infected with Rous sarcoma virus in vitro. Classical appearance is illustrated above. Virus is present at cell membrane surface or inside clear vacuoles (→). In figure 25, the "gray body" type of formation differs from usual empty vacuole, while in figure 20 :i totally different type of particle is demonstrated. Its relationship to the other is still unknown. X 42,000.


Discussion
Dr. Dalton (National Institutes of Health): Dr. Haguenau, what is your interpretation of the multivesicular bodies which you described? Do you consider them to be a fairly regularly occurring component in relation to virus infection?
Dr. Haguenau: It would appear that they are probably part of the Golgi apparatus in normal cells, but the ones seen in human breast cancer do not quite fit this description.
Dr. Dalton: Do you regard the presence of Rous virus particles in vacuoles as always the product of phagocytosis?
Dr. Haguenau: Yes, I think that those in true vacuoles have been engulfed by the cell rather than formed there in this particular material in vitro.
Dr. Gaylord (Yale University): I was amazed to see Dr. Haguenau's pictures of the structures in the Rous sarcoma cell nuclei. It is difficult to understand why they have not been seen before, since they are so conspicuous. Were those tumor cells or cultured cells?
Dr. Haguenau: The larger granules which I showed were in the cytoplasm and not in the nucleus. In the nucleus there were only small granules, perhaps in the range of size of polyoma virus. These are present in Rous tumor cells.
Dr. Gaylord: Were the patches of small particles numerous?
Dr. Haguenau: They were in cultured cells. The "dense bodies" have not been in vivo.
Dr. Fogh (New York State Health Department): It is sometimes difficult for a nonprofessional niorphologist to understand your criteria for designating a particle as virus. Is it ever possible from a purely morphologic observation to identify a particle as virus?
Dr. Haguenau: Yes. Some of the viruses have very definite features which distinguish them from any other type of particles. For instance, the avian viruses are all alike, and there is no doubt that they have a nucleoid and a double membrane and that there is no other structure in the cytoplasm of the normal cell which has the same aspect.
Dr. Howatson (Ontario Cancer Institute): I would like to ask Dr. Haguenau whether she has found in the polyoma virus any evidence of formation of the particles in the cytoplasm, particularly within mitochondria or bodies derived from mitochondria.
Dr. Haguenau: 1 have not really worked with this virus, but I know that Bernhard has seen particles in the cytoplasm. I have never seen a picture of his in which there appeared to be an actual site of formation in the cytoplasm. Have you?
Dr. Howatson: We think we have.
Dr. Parsons (Oak Ridge National Laboratory): I would like to confirm Dr. Haguenau's observations about the compound vacuoles, particularly in cancer tissue. In a study of 13 cases of acute leukemia and Hodgkin's disease, this was one of the most striking features of these tissues as compared with normal white cells. These particular structures, little vesicles inside large ones, which Law has called the compound vacuole, were very prominent, particularly in Ilodgkin's disease. It would appear that the term "multivesicular bodies" should be limited to primitive leukocytes. This was a term given by Porter to a very distinct and specific structure in leukocytes which, as far as I know, is not seen anywhere else.
Dr. Fogh: In an electron microscopic study made in collaboration with Dr. Edwards, in which we studied sections of normal amnion cells in the membrane, of primary amnion cells in primary culture, and of transformed amnion cells, we saw the multivesicular bodies only in the transformed cells.
Dr. Dalton: In your material in tissue culture, have you found an apparently definite life cycle of the Rous virus, as the group at Duke have shown in relation to the gray bodies in the development of myeloblastosis? In some material from one of Dr. Bryan's chickens that Dr. Ziegler has been looking at, we found particles in the lumen of the pancreas in young chicks. The sections showed a budding of these particles from the surface of the cell membrane just as one finds in other tumor cell types, for example, as with the Friend agent.
Do you feel that, in the tissue-culture studies on Rous virus, you are dealing with a special situation and a special environment, or do you think that the Rous agent is different from this unknown avian virus which we saw?
Dr. Haguenau: Do you really think that this is a typical Rous virus particle that you are referring to?
Dr. Dalton: I am only claiming it to be an avian virus, because it is in a chick.
Dr. Haguenau: I do not think that your pictures resemble the typical Rous virus particle. Your particle has no dense nucleoid or double membrane around it. That it seems to bud off is very unusual for the Rous virus, but may be related to the fact that you are dealing with an epithelial cell instead of a mesenchymal cell; that I do not know, but this story is not clear.
Dr. Beard: Dr. Dalton's picture is much like that seen in erythroblastosis. I would guess offhand that his particles were not Rous virus at all but erythroblastosis.
Dr. Banfield (National Institutes of Health): Dr. Haguenau, would you say more about the particles that you saw in your examinations of human breast cancer- where they occur, and their size? How often have you seen them and have you ever found dark bodies in the nuclei or anything that you might think were these dark bodies?
Dr. Haguenau: Yes, but this is a very infrequent finding. I have seen it about 3 times out of about 100 cases which we have examined. However, it was only by looking at photographs afterwards that I discovered them. I never looked for them on the microscope screen. It may well be that they are more frequent. Nevertheless, when they are seen, they are always in the nucleus and associated with the dense bodies. There are both the granular kind of structure and the dense bodies in the nucleolus, too, in human breast cancer.
Dr. Parsons: I would like to ask Dr. Haguenau her impression of the present status of osmic acid fixation, that is, whether she thinks the osmic acid gives a good appearance of the interphase mammalian nucleus or whether there is more that can be learned by some other staining technique. It seems that osmic acid is deficient in some way in staining the nucleus, particularly because of recent reports dealing with leukemic cells as having very abnormal chromosome configurations. If fixation could be improved, it might be possible to distinguish something definite between cancer cells and normal cells.
Dr. Haguenau: This is evident, and we would very much benefit by the use of other fixations for the study of nucleus, but at the same time, I do not feel that osmic acid fixation gives a distorted image. With our actual notion of the structure of the interphase nucleus, the idea of very thin coiled filaments cut tangentially in many sections does fit with the image we get.
Dr. Gey (Johns Hopkins Hospital): I would like to ask whether, in differentiating normal and tumor cells, you believe, as I do, that in the malignant cells, regardless of whether or not they are infected with viruses, one would find altered and decreased number of mitochondria and whether, in many malignant cells, there is not also a feedback mechanism involved in the uptake of materials from degenerating cells into competent cells which do take up these materials.
You mentioned something about some myelin figures and other structures. Do you not also identify many particles that have come from other cells?
Dr. Haguenau: Yes. Concerning mitochondria there is a definite answer. We have recently studied rather extensively the renal carcinoma of man, the clear-cell type, in which one of the most striking features is the amazing increase in mitochondria. Although the kidney tubule cell is full of mitochondria normally, the most striking feature in this growth is that nearly the whole cytoplasm is filled with mitochondria. Thus, instead of a decrease in number, usually considered particular to cancer, there are remarkably large numbers of mitochondria in this growth.
With respect to the turnover and engulfment of substance that might come from other cells, I am sure that this may occur, but I do not know of pictures that illustrate it.
Virus of Avian Myeloblastosis. XVII. Morphology of Progressive Virus-Myeloblast Interactions In Vitro
Continue to:
My Books

