The Fine Structure Of The Virus
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
The Fine Structure Of The Virus
High-magnification studies of the virus were facilitated by the lead hydroxide technique described by Watson (6). With this procedure a supplementary membrane of the virus located between the outer and the inner virus shells and measuring approximately 60 A in thickness was demonstrable. The mean dimensions of the virus are shown in the schematic drawing in text-figure 1.
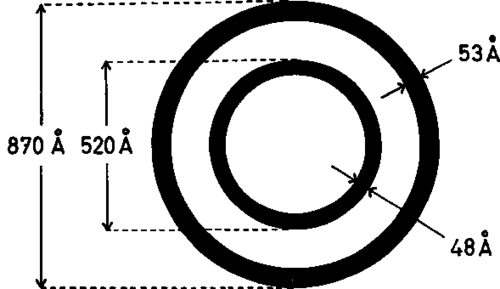
Text-figure 1. This schematic drawing illustrates only the mean dimensions of the shells of the Swiss mouse leukemia virus. These dimensions are subject to wide individual variations. No attempt has been made to indicate the precise distribution of the electron-dense material in this virus-membrane system and the high density of the inner virus shell observable after lead hydroxide staining.
The density of the virus material is usually higher in the peripheral portion between the outer and the inner shells than in the central portion of the virus. Such a virus may be related to the virus type "A" described by Bernhard (7). Occasionally, however, the reverse situation is observed, the center presenting the maximum electron density corresponding to virus type "C" described by Bernhard. In this case the dense material present in the center of the particle is usually irregularly distributed, without any suggestion of a twisted filament. In a few instances, however, a dense nucleoid is observed. The intermediate membrane is not always recognizable, and it is not yet clear whether it should be considered a definite component of the virus or whether it corresponds to the condensation of the material between the outer and the inner virus shells. The lead hydroxide technique does not seem to modify the dimensions of virus components and greatly increases the contrast of the virus, especially at the level of the inner shell which presents a maximum affinity for lead hydroxide. This remarkable affinity of the inner shell for the lead hydroxide is comparable in intensity to the affinity presented for the same salt by the cytoplasmic ribonucleoprotein (RNP) granules.
Relationship Of Virus To Cell Membranes And The Hypothesis Of Budding Of Virus From The Membranes
The frequency of the relationship between the viruses and cell membranes has been already demonstrated in the study of the virus fine structure. Further it has been possible to observe various forms of incomplete viruses appearing as a differentiation of the cell membrane itself. Since it is dangerous to describe dynamic processes from observations made on fixed tissues, it is fully recognized that the sequence of figures 1, 2, 3, and 4, in that order, is hypothetical. Nevertheless, by analogy with equivalent processes already known to occur in the development of other viruses, it is reasonable to consider those pictures as describing the different steps in maturation. The budding process seems to be the transitional phase of the virus cycle between the proviral stage and the mature extracellular infective particle. The rupture of the narrow pedicle probably liberates the mature virus into the intercellular spaces.
Two points more specifically related to present observations should be stressed: (1) Obviously the inner virus shell differentiates at the level of the cell membrane and does not correspond to a preformed vesicle coming from another area of the cytoplasm. (2) The virus, while budding from the cell surface, is definitely a type "A" particle with its maximum density at the periphery and not in the center. This indicates that the "A" particle is a recently formed virus and that the type "C" is possibly further along in development.
Continue to:
My Books

