Tumor Viruses. Speculation On Mechanisms. Part 2
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Tumor Viruses. Speculation On Mechanisms. Part 2
Some evidence for partial resistance of strain C57BL/Ka mice to the tumor-inducing effects of polyoma virus is cited. It is not known whether this presumed genetic influence is exerted directly at the cell-response level or indirectly at the immune-response level.
Organ-culture studies of the response to polyoma virus by submandibular gland of the mouse at different stages of morphologic and biologic development were reported. It was found that the proliferative response was most marked in adult glands and least marked and latest in appearing in 13- and 14-day rudiments of glands from embryos. In embryonic rudiments, cytolytic effects were noted first in the mesenchymal cells, then in epithelium. The morphologic character of the proliferative phase of the response differed for submandibular gland taken from an adult, immune female as compared with response of glands from newborn or embryo mice. In the former, there was wider spread of proliferating epithelium into the gelatin sponge, and a greater tendency toward a gland-forming pattern.
Our present impression, based on these still incomplete organ-culture studies, is that adult submandibular gland is more susceptible to the proliferation-inducing effects of polyoma virus than is the same tissue from newborn or embryonic mice. The advantage of newborn animals for tumor induction in vivo appears to depend on the overlap in time of (1) a period of weak immune response and (2) a period of intermediate capability for proliferative response.
A concept concerning intercellular relationships influencing the development of tumors in the salivary-gland/polyoma virus system is outlined. In this double-dependency scheme, a balance between virus and epithelial cell, favoring cell proliferation but restraining viral proliferation and resultant cytolysis, is maintained during the early stages of tumor genesis by the mesenchymal cells specifically responsible normally for morphologic and functional control. Investigations for testing this concept are now possible by techniques developed in the field of experimental embryology.
Plate 7
Figure 1. Low magnification showing 2 parotid-tumor nodules (dark areas at left and right) in which lymphoid cells make up majority of cell population. Note general resemblance of nodules to lymph nodes. Other tumor foci not exhibiting lymphocytic accumulations are also present. From a strain C3H/BiLw mouse inoculated within 24 hours after birth with polyoma virus, and autopsied when 3.5 months old. Hematoxylin and eosin. X 15.
Figure 2. Higher magnification of a parotid tumor of same type shown in figure 1. Tubular epithelial structures made up of cells with large, pale nuclei at right. Note varying stages of maturation of lymphoid cells, as in normal lymphocytogenesis. From a strain C3H/BiLw mouse inoculated within 24 hours after birth with polyoma virus, and autopsied when 3 months old. Hematoxylin and eosin. X 610.
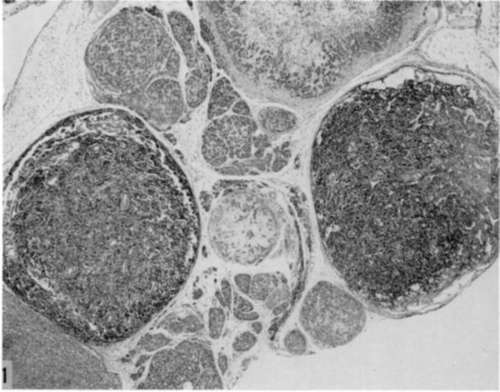

Plate 8
Figure 3. Lobule of extraorbital lacrimal gland partially replaced by polyoma virus-induced tumor (below, and center). A'ote large size of nuclei of non-tumor acinar cells in periphery of circumscribed lobule. Many giant mitotic figures are present among cells. Arrow (lower left) indicates mitotic figure in tumor cell. Hematoxylin and eosin. X 290.
Figure 4. Normal cxtraorbital lacrimal gland of young adult mouse. Compare nuclei of acinar cells with nuclei of similar cells adjacent to tumor focus in figure 3. Hematoxylin and eosin. X 290.


Plate 9
Figure 5. Giant mitotic figure (right center) in acinar cell of extraorbital lacrinal gland, adjacent to tumor focus (lower left). More than 100 chromosomes were estimated present in large mitotic figure. Telophase mitotic figure within polyoma virus-induced tumor is present Mow and to left. Hematoxylin and eosin. X 880.
Figure 6. Gelatin sponge matrix culture of submandibular salivary gland from a 5-month-old male C3H/Bi mouse, 30 days after explanation and exposure to polyoma virus. Solid clumps and cords of altered cells surround original explant (center) and extend into adjacent gelatin sponge. Hematoxylin and rosin. X 200.

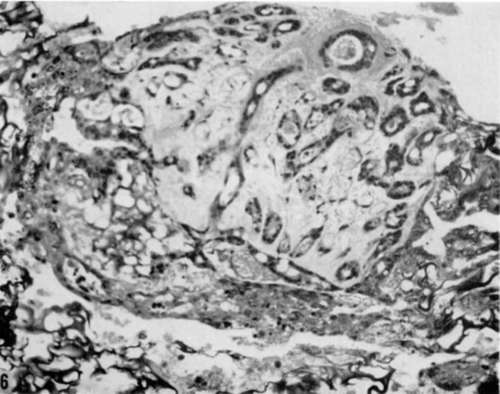
Plate 10
Figure 7. Gelatin sponge matrix culture of submandibular gland from 8-week-old C3H/Bi male mouse, 30 days after explantation and exposure to polyoma virus. Note extensive growth of altered epithelium into the sponge to right of explant. Compare with uninfected control culture in figure 8. Hematoxylin and eosin. X 130.
Figure 8. Gelatin sponge matrix culture of submandibular gland from same animal donating explant shown in figure 7. Explant shown here was also cultured for 30 days, but was not exposed to polyoma virus. Note absence of epithelial proliferation from margins of, or within explant. Also note many nuclei of viable stromal cells, as compared with their nearly complete absence in explant in figure 7. Hematoxylin and eosin. X 130.


Plate 11
Figure 9. Higher magnification of area shown in figure 8. Note persistence of normal branched glandular pattern and uniformity of cell nuclei. Compare with cellular changes in figure 10. Hematoxylin and eosin. X 780.
Figure 10. Higher magnification of area shown in figure 7. Note large nuclei and nucleoli, and basophilic cytoplasm of altered epithelium growing within compartments of gelatin sponge. At right center is mitotic figure close to nuclear remains of disintegrated cells. Hematoxylin and eosin. X 780.
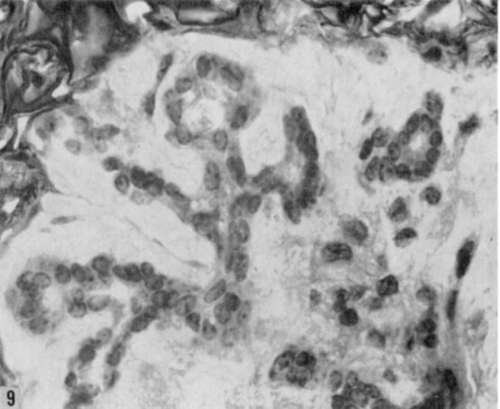
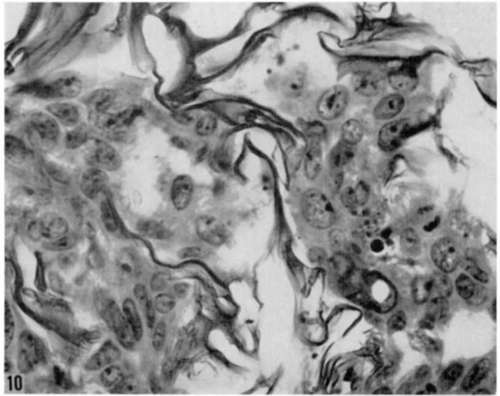
Plate 12
Figure 11. Section of tumor of salivary gland in a C3H. Bi mouse, nearly 12 months after mouse received subcutaneous inoculation of polyoma virus from tenth serial passage in P388 D1 cell cultures. Hematoxylin and eosin. X 290.
Figure 12. Section of tumor that grew subeutaneously over lumbar spine, at site of transplantation of 30-day culture of adult C3H/Bi submandibular gland infected with polyoma virus from tenth serial passage in P388 D1 cell cultures. Note marked similarity of cellular pattern here to that of tumor induced in riro, shown in figure II. Hematoxylin and eosin. X 290.
Figure 13. Section of polyoma virus-induced tumor of salivary gland, in its first transplant generation. Perivascular arrangement of cells is less pronounced, but general features are similar to those in tumor in figures 11 and 12. Section was from a transplanted parotid tumor described by Law, Dunn, and Boyle and was illustrated in their figure 24, plate 39. It was induced by direct extract from leukemic tissue. Hematoxylin and eosin. X 290.

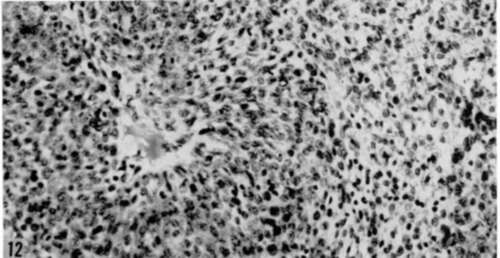

Plate 13
Figure 14. Gelatin sponge matrix culture of submandibular gland from 8-month-old C3H/Bi female, with HI titer of 1:800. After 30 days in culture, unexposed to polyoma virus in vitro. Note, preservation of glandular pattern and lack of growth or migration of epithelium into adjacent sponge. Hematoxylin and eosin. X 260.
Figure15. Culture from same donor and of same duration as that shown in figure 14, but infected with polyoma virus at time of explanation. Note glandular pattern of epithelial growth extending widely into sponge. Actual extent of growth away from explant (left) was more than twice the distance shown here in all directions. Compare with figure 14. Hematoxylin and eosin. X 260.


Plate 14
Figure 16. Another area from same culture shown in figure 15 to demonstrate continuity of epithelium of original explant (center) with altered epithelium extending centrifugally into sponge. Hematoxylin and eosin. X 340.
Figure 17. Section of duplicate culture to that shown in figures 15 and 16.Note gland-like pattern of altered epithelium growing within compartments of sponge at distance from original explant (not shown). Mitotic figure at upper left. Dark basophilic body within lumen at lower right is pyknotic nucleus of dead cell. Hematoxylin and eosin. 570.


Continue to:
My Books

