Rhythmic Segmentation Of The Intestinal Contents
Description
This section is from the book "The A. B. - Z. Of Our Own Nutrition", by Horace Fletcher. Also available from Amazon: The A. B.-Z. Of Our Own Nutrition.
Rhythmic Segmentation Of The Intestinal Contents
This is by far the most common and the most interesting mechanical process to be seen in the small intestine. The nature of the process may best be understood by referring to the diagram in Figure 2. A string-like mass of food is seen lying quietly in one of the intestinal loops (line I, Fig. 2). Sud denly an undefined activity appears in the mass and a moment later constrictions at regular intervals along its length cut it into little ovoid pieces. The solid string is thus quickly transformed, by a simultaneous sectioning, into a series of uniform segments. A moment later each of these segments is divided into two partides, and immediately after the division neighbouring particles (as a and b, line 2, Fig. 2) rush together, often with the rapidity of flying shuttles, and merge to form new segments (as c, line 3, Fig. 2). The next moment these new segments are divided, and neighbouring particles unite to make a third series, and so on. At the time of the second segmentation (line 3, Fig. 2) the end particles are left small.
Observation shows that these small pieces are not re-divided. The end piece at A simply varies in size with each division; at one moment it is left small, at the next moment it is full size from the addition of a part of the nearest segment, and a moment later is the small bit left after another division. The end piece at B (probably the rear of the mass) shoots away when the end mass is divided, and is swept back at each reunion to make the large end mass again, only to be shot away and swept onward with each recurrence of the constrictions. Thus the process of repeated segmentation continues, with the little particles flitting towards each other and the larger segments shifting to and fro, commonly for more than half an hour without cessation. From the beginning to the end of a period of segmentation the food is seen to have changed its position in the abdomen to only a slight extent; whether this change is a passing of the food along the loop, or a movement of the loop itself, it is impossible to tell from the shadows on the screen. The change of position, however, is much less conspicuous than the lively division and redivision which the mass suffers so many times from the busy, shifting constrictions. From this typical form of rhythmic segmentation there are several variations.
Sometimes, and especially when the mass of food is thick, the constrictions do not make complete divisions and are so far apart that the intermediate portions are relatively large. Moreover, the constrictions do not take place in the middle of each portion, but near one end; thus each portion is constricted, not into halves, but into thirds. If a little pointer is placed at the middle of a segment, when the segments are completely divided into halves, in a few seconds the pointer will be in the middle of the clear space between two segments; but in a few seconds more the first phase will return and the pointer will again be indicating a segment, - two operations intervene between similar phases. When, however, the portions are constricted into thirds, the indicator shows it, since three operations intervene between similar phases. The manner of these changes is made clearer by reference to the diagram in Figure 3. That each portion is constricted into three pieces is proved also by watching the gradual reduction of the portion at the left end of line 1 through lines 2 and 3, and also in the gradual formation of a full-sized portion at the right end of lines 2, 3, and 4. When food undergoing this process is watched, it appears to be affected by a series of constrictions, each of which starts at one end of the mass and marches through to the other end, leaving its impress at short intervals along the length.
The progression of the dotted lines from right to left in a, b, c, and d, etc., Fig. 3, gives a notion of these advancing constrictions. Another variation of the segmentation is shown in Figure 4. In this type there are evidently divisions and subdivisions, i. e., one more operation between the appearance and the reappearance of the same phase than is present in the simple division of the small segments in a long string of food (Fig. 2 ). This form of segmentation is fairly typical for the constrictions seen in food advancing through the intestine. Sometimes the divisions occur in the middle of a long string of food and leave the ends wholly unaffected.
Figure 1. - Appearance of food in the intestines 5 3/4 hours after eating. This and other radiographs reduced two-thirds.

Figure 2. - Diagram representing the process of rhythmic segmentation. Lines 1, 2, 3, 4 indicate the sequence of appearances in the loop. The dotted lines mark the regions of division. The arrows show the relation of the particles to the segments they subsequently form.
A remarkable feature in the segmentation of the food is the rapidity with which the changes take place. The simplest way of estimating the rate of division is to count, not the number of times the partition of the food recurs in the same place, but the number of different sets of segments observed in a given period. Thus in Figure 4 the appearances of lines 1, 2, 3, 4, etc., would be counted, and not merely lines 1, 4, etc. Repeated observations on different animals have shown that the most common rate of division in long, thin chains of food varies between twenty-eight and thirty times in a minute; i. e., there is a change from one set of segments to another set every two seconds, and a return of the same phase every four seconds. In some cases the rate is as low as twenty-three times per minute. The larger masses seem to be associated with a slower segmentation; the operations indicated in Figure 3, for example, occurred from eighteen to twenty-one times in a minute, so that the same phase reappeared only once in eight or nine seconds. The segmentation frequently continues for more than half an hour; in one instance it was seen to persist with only three short periods of inactivity for two hours and twenty-two minutes.
At the rate of thirty segmentations per minute it is clear that a slender string of food may commonly undergo division into small particles more than a thousand times while scarcely changing its position in the intestine.
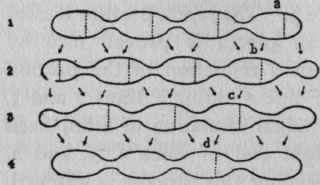
Figure 3. - Diagram showing the relations of the portions when they are constricted into three pieces. The dotted lines indicate regions of constriction; the arrows indicate the relationship of the pieces to the portions they subsequently form.
I have seen once, in a cat only lightly etherised, the exterior of an intestine which was dividing the food as above described. An hour and a half after a meal of salmon the anaesthetic was given, the abdomen opened, and the flaps raised so as to form walls. Warm salt solution was then poured into the abdominal cavity, and the floating coils left covered with the transparent omentum. The gastric peristaltic waves were running regularly; on the intestine there were visible at various places during the period of observation regions of constriction which had the appearance shown in Figure 3, except that the rings were relatively nearer together. New rings of constriction took place on the same side of all the bulging parts at the margin of the constricted portion (cf. dotted lines, Fig. 3). As new rings occurred the old relaxed, but apparently with tardiness, for the contents gurgled as if forced through the narrowed lumen. The constrictions recurred irregularly and at much longer intervals than in the normal animal.
The contracted rings were pale and bloodless.
The effect of the process of rhythmic segmentation proves it an admirable mechanism. The food over and over again is brought into closest contact with the intestinal walls by the swift kneading movement of the muscles. Thereby not only is the undigested food intimately mixed with the digestive juices, but the digested food is thoroughly exposed to the organs of absorption. Mall has shown that contraction of the intestinal wall has the effect of pumping the blood from the submucous venous plexus into the radicles of the superior mese-teric vein, and thus materially aids the intestinal circulation. Moreover, lacteals loaded with fat will in a few moments become empty unless the intestine is slit lengthwise, so that the muscles cannot exert compression. The rhythmic constrictions, therefore, both propel the blood in the portal circulation and act like a heart in promoting the flow of lymph in the lacteals. This single movement with its several results is an excellent example of bodily economy; the repeated constrictions, as already shown, thoroughly churn the food and digestive fluids together, and also plunge the absorbing mucosa into the very midst of the food masses: but not only are the processes of digestion and absorption favoured by these movements; they also, by compression of the veins and lacteals of the intestinal wall, serve to deport through blood and lymph channels the digested and absorbed material.
Continue to:
My Books

