Starvation. Part 11
Description
This section is from the book "The Elements Of The Science Of Nutrition", by Graham Lusk. Also available from Amazon: The Elements of the Science of Nutrition.
Starvation. Part 11
Frentzel1 has shown the effect of external work upon the protein metabolism of fasting dogs. One of the dogs did an amount of work corresponding to 216,937 kilogrammeters in three days. The protein metabolism rose during the working hours and continued high on the last day, which was one of complete rest. Frentzel computes that the nitrogen elimination of the last four days (= 20.7 grams) represents an energy equivalent of 220,300 kilogrammeters. This could not cover the work done by the dog if we add to the measured work that which was done by the heart and respiratory muscles. The protein metabolism of four days is therefore entirely insufficient to cover the work done during three. The source of the energy for the work accomplished must be found in an increased metabolism of fat. The increase in protein metabolism above that of rest was not sufficient to supply 7 per cent. of the energy needed to do the work. The record of the dog's nitrogen metabolism is as follows:
Influence Of Work On The N Metabolism Of Fasting Dogs
Day. | Work or Rest. | Food. | Grams of N Excreted. | |
Per Day. | Per Hour. | |||
1st to 4th...................... | Rest. | 100 g. lard | ||
5th............................ | Rest. | 100 g. " | 3.13 | 0.1304 |
6th............................ | Rest. | 100 g. " | 3.52 | 0.1467 |
7th............................ | Rest. | Fasting. | 3.71 | 0.1546 |
8th............................ | Rest. | " | 3.99 | 0.1663 |
9th............................ | Work. | " | 4.97 | 0.3680* |
0.1837! | ||||
10th............................ | Work. | " | 5.02 | 0.2750* |
0.1960† | ||||
nth............................ | Work. | " | 5.03 | J 0.2400* |
0.2335† | ||||
12th............................ | Rest. | " | 5.08 | 0.2117 |
* Work.
†Rest.
Succi did not show a similar rise of protein metabolism from the effect of work. The eleventh day of his fast he spent in bed. On the twelfth day he rode a horse for an hour and forty minutes, raced for eight minutes with some students, and gave an exhibition of fencing in the evening. During the day he walked 19,900 steps. The urinary nitrogen on the eleventh day (rest) was 7.88 grams; on the twelfth (work), 7.16; and on the days following 3.50, 5.33, 5.14, 5.05. The work done was evidently at the expense of increased metabolism of fat. That this is the case had already been demonstrated by Pettenkofer and Voit.1 A fasting man at work showed no increase in his protein metabolism, but the quantity of fat burned rose enormously. This is shown by the following comparison of the number of grams of fat burned:
1 Frentzel: "Pfluger's Archiv," 1897, lxviii, 212.
Day | Night. | ||
8 a.m. to 8 P.M. | 8 p. m. to 8 a. m. | ||
Rest during day..................... | 116 gm. | 94 gm. | |
Work during nine hours of day period.. | 312 gm. | 70 gm. | |
The fat metabolism during the day of work is two-and-a-half times that of the resting day, and is presumably the source of the energy for the mechanical work accomplished. During the night following the working day the reduction of fat combustion as compared with the night before is due to more profound sleep.
Another phase of the effect of work is shown in the variation between the day and night metabolism of Tigerstedt's fasting medical student, J. A. The average carbon dioxid excretion in grams for two-hour periods during five days of fasting was as follows. The figures showing the elimination during the hours of sleep are printed in black:
A.M. | P.M. | ||||||||||
Time ..................... | 10-12 | 12-2 | 2-4 | 4-6 | 6-8 | 8-10 | 10-12 | ||||
Carbon dioxid (grams)..... | 54.8 | 57.2 | 54.1 | 57.8 | 59.5 | 66.4 | 46.5 | ||||
A.M. | |||||||||||
Time............................ | 12-2 | 2-4 | 4-6 | 6-8 | |||||||
Carbon dioxid (grams) ................. | 37.5 | 39.1 | 40.7 | 68.6 | |||||||
1 Pettenkofer and Voit: "Zeitschrift fur Biologie," 1866, ii, 459; C. Voit: Ibid., 1878, xiv, 144.
The nitrogen of the urine was also less during sleep than during the waking hours:
Fasting Day. | N in the Urine. | |
Day. | Night (10 p. m. to 10 A. M.). | |
ISt................................... | 7.11 | 4.93 |
2d.................................... | 6.87 | 5.85 |
3d.................................... | 6.83 | 6.65 |
.4th................................... | 7.91 | 5.65 |
5th................................... | 6.36 | 4.98 |
Johansson1 finds that the inequality of night and day metabolism depends on muscular work. Sitting up raises the metabolism, and standing does so still more. Even when one lies in bed, restlessness during the day may increase the metabolism. Zuntz2 was the first to mention the condition of absolute muscular rest as significant. Even when perfect muscular relaxation ensues there may still be influences, such as light on the retina or sounds, which may act reflexly on the organism and slightly increase the metabolism. Johansson illustrated these variations in the following comparisons between night and day excretion of carbon dioxid of starving men, the night C02 being figured at 100:
Night CO2. | Day CO2. | Author. | |
Complete muscular rest........... | 100 | 105 | Johansson. |
Ordinary rest in bed.............. | 100 | 110 | Johansson. |
Ordinary life (no hard work)...... | 100 | 142 | Tigerstedt. |
" " " " | 100 | 128 | Pettenkofer and Voit |
" " " " | 100 | 147 | Tigerstedt. |
Johansson agrees with Tigerstedt that the minimum metabolism of a man in bed is represented by 24 to 25 calories per kilogram daily, and results obtained by Zuntz, Loewy, and others lead to the same conclusion.3
1Johansson: "Skan. Archiv fur Physiologie," 1898, viii, 109.
2 Lehmann and Zuntz: "Virchow's Archiv," 1893, cxxxi, Supplement, 26.
3 Tigerstedt: "Skan. Archiv fur Physiologie," 1910, xxiii, 302.
The temperature of the fasting organism is usually normal. Luciani found a normal temperature in Succi during his thirtyday fast. The temperature falls only a few days before death. Sonden and Tigerstedt1 find that the diurnal variations persist during fasting in their ordinary rhythm. The average temperature of the medical student J. A. during his five-day fast was but 0.16 degree below his normal temperature when food was allowed him. These diurnal variations are exactly concomitant with the fluctuations of carbon dioxid excretion noted on a previous page. When the carbon dioxid production increases, the temperature rises.
This parallelism may be easily shown by comparing the two factors in the chart (Fig. 5) as given by Sonden and Tigerstedt.1 Furthermore, the diurnal variations tend to disappear if the person be kept in a state of muscular rest, so that the output of energy during the day and the night remains the same. In this state the temperature may fall 0.6 degree below the normal on account of the absence of muscle movement. This regularity of temperature and metabolism is beautifully shown in Fig. 6 taken from Johansson.1

Fig. 5. - Curve of carbon dioxid elimination compared with Jurgensen's curve of normal diurnal temperature variation. This individual led a normal life and partook of his usual nourishment.
1 Sonden and Tigerstedt: "Skan. Archiv fur Physiologie," 1895, vi, 136.
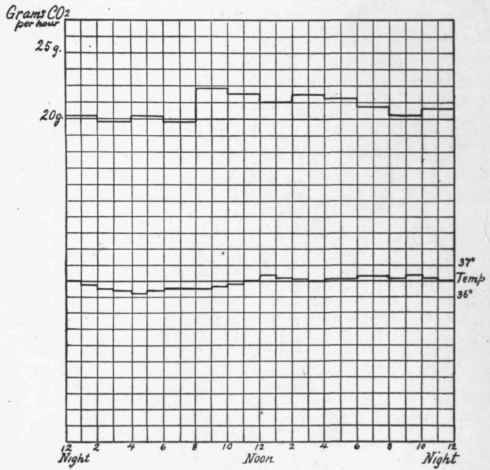
Fig. 6. - Carbon dioxid elimination and body temperature in fasting and complete muscular rest.
1 Sonden and Tigerstedt: Ibid., p. 132.
Inversion of the normal routine of life, so that one sleeps in the daytime and is awake and active at night, brings about an inversion of the curve of body temperature. This is well shown in the monkey.2
Benedict,3 however, was unable to obtain any inversion of the curve of normal body temperature in men who worked during the night and slept during the day.
Gibson4 traveled half-way round the world in making a trip from New Haven, Connecticut, to Manila, and then returned. He found that the rhythm of daily variation was dependent on the time of the solar day and was independent of the part of the world in which he happened to be.
1 Johansson: Loc. cit., p. 142.
2 Galbraith and Simpson: Proceedings of the Physiological Society, "Jour, of Phys.," 1904, xxx, p. xx.
3 Benedict: " Amer. Jour. of Phys.," 1904, xi, 145.
4 Gibson: "Amer. Jour. of Med. Sci.," 1905, cxxix, 1049.
Continue to:
My Books

