Chapter IV. The Regulation Of Temperature
Description
This section is from the book "The Elements Of The Science Of Nutrition", by Graham Lusk. Also available from Amazon: The Elements of the Science of Nutrition.
Chapter IV. The Regulation Of Temperature
It has been seen that the temperature of a warm-blooded animal is maintained at the normal throughout a, fast. Not only this, but it is maintained at the same level, even though the temperature of the outside environment vary from o˚ C. and lower to 30˚ to 35˚ C. In cold-blooded animals the temperature of the body is only slightly higher than that of their environment at the time. The metabolism of such animals varies with the temperature. The frog in the mud during the winter at a temperature of 4˚ C. has quite a different metabolism from that which he enjoys during the summer sunshine as he sits on the river-bank or snaps at passing flies. The curve of his carbon dioxid elimination at various temperatures has been made by E. Voit from the analyses of H. Schulz,1 and is given below:
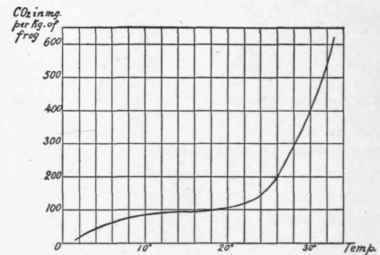
Fig. 7. - CO2 in milligrams per hour per kg. frog.
Krogh2 finds that the rise in the metabolism of the normal frog which appears at 20˚ C. does not show as sharp an ascent in narcotized animals, indicating that in the normal frog nervous influences which produce tone begin to make themselves felt at this temperature.
1 Schulz: "Pfluger's Archiv," 1877, xiv, 78.
2 Krogh: "Internat. Zeitschr. f. physik.-chem. Biologie," 1914, I, 492.
Rohrig and Zuntz1 first showed that a curarized mammal at ordinary room temperature lost the power of maintaining its body temperature, and the intensity of its metabolism decreased accordingly. Curare prevents the transmission of motor impulses to voluntary muscles.
Krogh states that the curve of oxygen absorption as influenced by body temperature is the same in the anesthetized frog and fish as in the curarized dog. One of Krogh's curves which is given here shows a sixfold increase of oxygen absorption in the curarized dog, indicated by a rise from 2.1 c.c. per minute at a body temperature of 140 C. to 13 c.c. per minute when the body temperature reached 37.20 C.
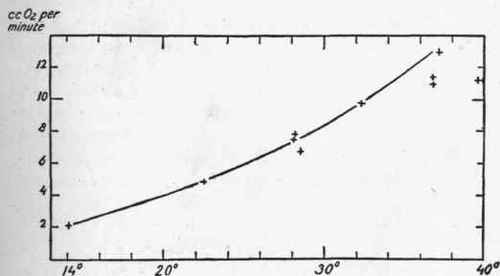
Fig. 8. - Curve of metabolism of a curarized dog subjected to different temperatures (after Krogh).
If the sciatic nerves of a curarized dog be severed, Mansfield and Lukacs2 found that the heat production falls 10 or 15 per cent., but if the sympathetic nerves had previously been severed, cutting the sciatic was without influence upon metabolism. From this they conclude that in the curarized animal sympathetic nerves carry impulses which produce tone in muscles.
1 Rohrig and Zuntz: "Pfluger's Archiv," 1871, iv, 57.
2 Mansfield and Lukacs: "Pfluger's Archiv," 1915, clxi, 467.
The reduction in activity which accompanies reduced body temperature is exemplified by the fact that a cat whose temperature has been artificially reduced to 19° C. may have but one heart-beat per minute.1 At the time of hibernation the marmot lives at the expense of fat. The metabolism may correspond to only one-thirtieth the amount of energy used during the period of activity.2
Hehriques3 reports concerning the metabolism of a hedgehog awakening from the winter sleep. The animal weighed 660 grams.. Tracheotomy was performed and the respiratory exchange determined during half-hour periods with five-minute intervals between periods, by the Zuntz method. The animal's body temperature was 6.50 C. in the rectum at the start, and the room temperature was 130 C. The animal remained quiet except for characteristic muscle movements resembling shivering which always accompany the awakening of hibernating animals. The results were as follows:
Period. | I. | 11. | 111. | IV. | V. |
R.Q.................... | 0.62 | 0.70 | 0.72 | 0.71 | 0.70 |
Oxygen, c.c, hour and kg.. | 375 | 334 | 851 | 1983 | 2083 |
Rectal temperature at end | 7.5º | 9.0º | 10.70 | 14.1º | 26.6º C. |
Contrary to other authors, Henriques concludes that the awakening from winter sleep is usually at the expense of fat and not of carbohydrate.
During the entire period 2.21 liters of oxygen were absorbed by the animal. If this had been used for the oxidation of fat the heat produced would have been 10.40 calories (2.21 X 4.7 cals., see p. 61). At the conclusion of the experiment the animal was killed and placed in an ice calorimeter. It was found that the whole animal gave off 13.41 calories. At the beginning of the experiment when the rectal temperature was 6.50 C. the hedgehog contained 3.56 calories (6.50 C. X 660 grams X 0.83 factor for specific heat of body). Therefore 9.85 calories were added to the body during the period of awakening, and these could have been obtained from the 10.40 calories derived from oxidation of fat and still leave a surplus of 0.55 calorie for loss of heat through radiation and conduction during the period when the body temperature was above the temperature of the environment. It should be remembered, however, that during the earlier periods of low body temperature the organism must have gained heat from its environment.
1 Simpson and Herring: "Journal of Physiology," 1905, xxxii, 305.
2 Regnault and Reiset: "Annates de chem. et de physic," 1849, xxvi, 299.
3 Henriques: "Skan. Archiv f. Physiol.," 1911, xxv, 15.
E. Voit 1 has drawn attention to the fact that a curve of increasing metabolism with increasing temperature corresponds to the increasing ability of muscle to contract, and to the increasing effectiveness of enzymotic activity. A high temperature is necessary for the irritability and activity of protoplasm. The warmth of the sunshine increases the irritability of the protoplasm of the tree in the spring, with the resulting development of the foliage. Warmth, however, is no not the cause of the metabolism, but only one of the conditions for it. In warm-blooded animals the temperature is mamtained at a constant level independent of climatic conditions, and this level is a favorable one for the activity of nerve and muscle. It would indeed be inconvenient were the active life of a man dependent upon the temperature of his environment. The essential mechanism for the regulation of the body temperature is nervous. The action of cold on the skin may stimulate its peripheral nerve-endings, which are sensitive to cold, and reflexly effect in the organism a greater heat production and a vasoconstriction of peripheral blood-vessels; the action of heat, on the contrary, effects vasodilatation and production of sweat. It is believed that the cold-blooded progenitors of warm-blooded animals changed their habitat from the sea to the land at a tropical temperature which is at present possessed by their descendants. In the course of development these animals acquired the power to maintain that ancestral temperature which proved favorable for the activity of their body substance. The nervous mechanism through which this is accomplished is twofold: First, there is an increased production of heat in the presence of external cold (the chemical regulation of temperature); and, second, variations in the quantity of blood supplied to the skin modify loss of heat by radiation and conduction, and variations in the amount of sweat modify the loss of heat by evaporation of water (these are the factors of the physical regulation of temperature). The great importance of these two controlling influences will be seen as the subject develops.
1E. Voit: "Sitzungsber. der Ges. fur Morph. und Physiol.," 1896, Heft HI.
Continue to:
My Books

