Localization Of The Cerebral Functions
Description
This section is from the book "A Manual Of Physiology", by Gerald F. Yeo. Also available from Amazon: Manual Of Physiology.
Localization Of The Cerebral Functions
Whether the entire surface of the hemispheres can be mapped out into areas, each of which is set apart for a definite immutable function, is a question surrounded with difficulty, and which, up to the present, cannot be answered with certainty. The experimental evidence hitherto brought forward on the subject seems, in some points, to be contradictory, a fact which may be explained partly by the difficulties with which such experiments are beset, and partly by different observers being anxious to uphold with too great fervor either the localization or non-localization theory.
The evidence on this subject may be briefly summarized as follows. In favor of localization are the facts that -
1. Lesion of a certain part of the cortex of the frontal lobe of the left hemisphere of man (posterior part of the third frontal convolution) has been so frequently followed by the loss of memory of words necessary for the faculty of speech - aphasia - that pathologists now call that spot the speech centre.
2. Destruction of the cortical surface of the angular gyri, or of the posterior lobes, causes transitory loss of the perception of visual impressions, and if an area including both angular gyri and posterior lobes be destroyed, the result is permanent blindness.
3. Destruction of the convolutions around and in the neighborhood of the fissure of Rolando gives rise to loss of power in the limbs of the other side, voluntary motion being abolished when an extensive area is destroyed. This loss of power is more obvious in animals with complex brains (man and monkey) than in those less highly organized (dog, cat, rabbit), which gradually recover.
4. The function of the part destroyed may be lost forever, and the nerve channels, which formerly carried the impulses to or from the injured centre, become degenerated.
5. Stimulation of the convolutions around the fissure of Rolando gives rise to definite coordinated movements of muscles of the other side. Local groups of muscle respond with striking constancy to the electric stimulation of certain definite and limited areas of the cortex. These convolutions have been mapped out into motor centres for hind limb, fore limb, face, etc. Compare Figs. 258 and 259.
From the foregoing Ave may safely conclude (1) that certain parts of the cortex of the hemispheres are the agents for the reception of definite sensory impressions; (2) that others (the neighborhood of the fissure of Rolando) are related to the discharge of voluntary motor impulses; and (3) though we cannot say that the anterior lobes are immediately subservient to either the sensory or motor functions, a portion of one of them seems devoted to the memory of words.
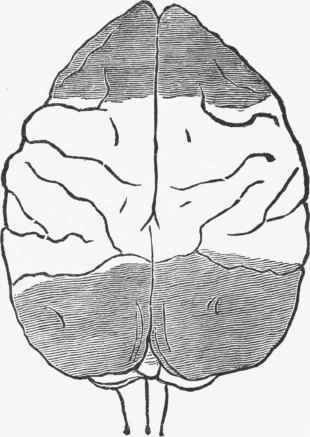
Fig. 262. The dark shading shows areas which were destroyed in a monkey without giving rise to any functional defect that could be detected. (Ferrier).
As objections to the soundness of these conclusions, it has been urged: -
1. Considerable discordance still exists in the results arrived at by different experimenters.
2. The function returns after the lapse of a variable interval, particularly in unilateral destruction of the cortical centres. In some instances the loss of function only continues for a few hours after the operation; in others (those in which the injury is extensive and deep, or the animal belongs to a class with high mental organization) the recovery is slow and may extend over several weeks or months.
3. Certain tracts of the cortex of the hemispheres, notably the anterior and posterior lobes, may be extensively injured, by accident or experiment, without interfering with the cerebral functions in any marked or tangible way. Both men and animals have lived for years after the loss of a considerable quantity of brain substance, without showing impairment of either mental or bodily faculties.
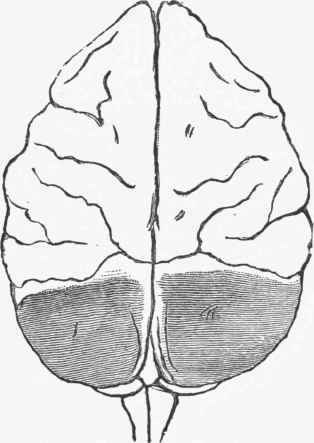
Fig. 263. Extirpation of the posterior lobes. Lesion followed by only transient disturbance of vision. In a few hours no defect could be detected in any of the animal's functions. Some observers have found total blindness after a lesion but slightly greater than that here shown. (Ferrier).
4. Certain areas of the brain surface may be stimulated mechanically, chemically, or electrically, without the least response being shown by the animal, to indicate either sensory or motor excitations.
In spite of this negative evidence from the facts just adduced - viz., that certain groups of muscles respond regularly to the stimulation of local areas of the brain surface, and that loss of function of some organ occurs when a given point is injured - it seems definitely fixed that certain local parts of the brain surface are in more immediate connection with definite peripheral organs than with others, and that these local areas have been in the habit of receiving or sending out special impulses.
The evidence is strongest in support of the motor areas situated around the fissure of Rolando in the central region of the hemispheres. Here (i) limited stimulus excites definite action, (2) circumscribed lesion is followed by local paralysis, and (3) after lesion a tract of degeneration unites the cortical and spinal centres engaged in the production of voluntary movement.
With regard to the sensory centres the areas are not so perfectly localized. We can hardly suppose that all the gray matter of the angular gyri and occipital lobes are devoted exclusively to the reception of visual impressions. Yet it has been stated all this region must be destroyed to annihilate the visual function.
From the fact that its stimulation causes movements of the pinna and its destruction is said to abolish hearing, the superior temporo-sphenoidal convolution has been allotted the function of an auditory centre. From somewhat similar evidence, and that gained from anatomy, the hippocampal lobule is said to be the seat of smell.
Ordinary sensation has been localized to the inferior temporo-sphenoidal convolution and the hippocampal region, owing to the anaesthesia found after destruction of these parts.
From the other facts mentioned - viz., the absence of functional disturbance after cortical lesion and the recovery of function after injury, we must conclude that there are extensive tracts containing cells to which we can assign no localized function, and that the local areas to which a function can be assigned are not the only agents which can carry on the business of receiving impulses from the periphery, and sending voluntary impulses to the muscles; but that there are many groups of nerve cells which can take on the duty of the injured cells and act for them in receiving sensory and discharging motor impulses.
It has already been pointed out that the function of any given nerve fibre depends on the relationship of its terminals. The fibre itself is merely a conducting agent. In somewhat the same way the functions of any given nerve cell must depend on the number and character of its connections. If it be connected with a motorial end plate in a muscle, it can only excite impulses that give rise to motion : if it be connected with a sensory terminal, it can only be a receiver of sensory impulses. But, in the gray matter of the spinal cord, and still more so in that of the cerebral cortex, we may assume that all the cells are in more or less intimate connection with innumerable other cells. In fact, we must imagine that the gray matter of both cord and brain is interwoven into a complex texture of fibrils and cells, no part of which is isolated from the rest, but all the elements form part of a continuous system, and within certain limits can subsidize each other's functions.

Fig. 264. The shading shows the great extent of surface destroyed before permanent cortical blindness followed by retinal atrophy was produced; i.e. all posterior lobes and angular gyri.
When we excise, cauterize or stimulate a given point of the complex cortex, we do not know in what way we interfere with the perfect action of that wonderful nervous nexus which controls the organism, for we can only judge of the effects we produce by results limited to those few functions the activity of which is obvious.
Continue to:
My Books

