The Spinal Cord. Continued
Description
This section is from the book "A Manual Of Physiology", by Gerald F. Yeo. Also available from Amazon: Manual Of Physiology.
The Spinal Cord. Continued
2. The crossed or lateral pyramidal tracts (Fig. 246, P) are continuous with the pyramids of the opposite side of the medulla where the crossing of the fibres is completed. They lie in the lateral white columns, occupying their posterior part next to the posterior gray columns, and are separated from the surface by the direct cerebellar tract (dc). The crossed pyramidal tracts also taper toward the lower part of the cord, but can be traced to the lumbar region. The fibres are connected with the anterior gray column of the same side.
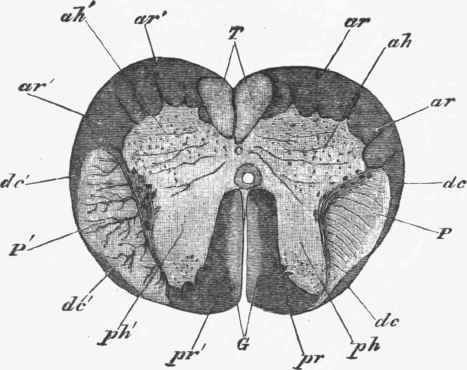
Fig. 246. Transverse section of the spinal cord of embryo at five months.
G. Columns of Goll. T. Direct pyramidal tracts. P. Crossed pyramidal tracts, dc. Direct cerebellar tract; ar. Anterior root zones; pr. Posterior root zones; ah. Anterior gray column; ph, Posterior gray column.
Section of these tracts in the cord of the pyramids in the medulla, or the channels leading from the motor areas, as well as destruction of the motor areas in the cortex of the brain, is followed by descending degeneration of both these pyramidal white columns along their entire extent. The track of degeneration corresponds with the limits assigned by developmental research to these pyramidal tracts. It therefore seems clear that they are the channels by which the efferent impulses from the brain travel to the cells of the anterior cellular column of the spinal cord. There seems to be no functional difference between those fibres that cross in the medulla at the decussation of the pyramids and those which pass directly from the medulla to the same side of the cord, and then gradually cross on their way to their destination.
3. The direct cerebellar tracts (dc) can be traced from the inferior peduncle of the cerebellum to the superficial part of the lateral white columns of the cord, where they form a flattened band of fibres which covers in the crossed pyramidal tract. The fibres appear to be connected with the cells of Clarke's column, to be described below. This tract tapers toward the lumbar region, about the upper limit of which it disappears.
After section of the cord ascending degeneration can be traced along these tracts through the restiform bodies of the medulla oblongata to the vermiform process of the cerebellum. Hence we may conclude that they carry centripetal impulses.
4. The posterior median columns (Goll) are thick strands of white fibres, triangular in section (Fig. 246, G), which form the immediate boundary of the posterior median fissure. They can be traced from the medulla to the mid-dorsal region, where they taper to a point. Some of the fibres of the posterior roots are connected with this column.
In Coil's posterior median column ascending degeneration can be traced, after section, up to the clavate nucleus of the medulla. This tract is therefore also afferent.
5. Anterior root zone (ar) is the name given to the white substance next to the anterior gray columns not included in the parts just described. These tracts do not taper toward the lower part of the cord, but vary in thickness with that of the roots of the spinal nerves, being thickest at the cervical and lumbar enlargements.
After transverse section of the cord descending sclerosis can be traced a short distance down these tracts. The direction of the degeneration shows them to be efferent, but from the limited extent of the sclerosis we may suppose that they only carry impulses from the upper to the lower regions of the spinal cord.
6. Posterior root zones (Burdach) (pr) surround the part of the gray column from which the posterior roots spring, and vary in proportion to the size of the roots of the spinal nerves.

Fig. 247. Diagram of transverse section of the cervical parts of the spinal cord, showing the white tracts supposed to be functionally distinct by differences of shading.
A. Anterior; P. Posterior median fissures, dp. Direct pyramidal; cp. Crossed pyramidal tracts; dc. Direct cerebellar; pm. Posterior median column (Goll); az. Anterior; pz. Posterior root zones.
The mode of degeneration (limited ascending sclerosis) of these fibres teaches us that they are afferent channels probably carrying impulses from the cells of the lower parts of the posterior gray columns to those in its upper segments.
Experimental Methods
Besides the foregoing anatomical facts, we learn from experimental research certain facts concerning the loss of function which follows transverse sections of different extent, of the spinal cord.
1. Complete section of the cord is followed by loss of sensation and the power of voluntary motion in the parts below the point of section (Galen).
2. Section of one side of the cord is followed by loss of sensation on the side of the body opposite to and below the section, and loss of all voluntary motion of the parts below and on the same side as the injury of the cord. Increased sensitiveness of the parts where the motor paralysis exists is also said to be observed in some of the lower animals.
3. If the gray matter and the anterior and posterior white columns of one-half of the cord be cut across, i. e., when only the lateral white columns remain intact on that side, both motor and sensory impulses (as observed in the rabbit) seem to be transmitted normally.
4. Longitudinal section of the commissures which unite the two sides of the cord, so as to separate the lateral halves, is said not to influence voluntary motion, but produce an ill-defined loss of sensation below the lesion.
5. Experiments consisting of partial and local sections were conducted with the object of determining the exact course of the impulses giving rise to different kinds of sensation; and it was concluded that ordinary sensory impulses (pain) traveled by the gray matter, while tactile temperature and muscle sense traveled via the posterior white columns. Though pathological observation and the occurrence of "analgesia" (unimpaired tactile sense with local loss of painful impressions) suggest the idea of such distinct paths for different kinds of sensation, it would appear that the localization of pain to the gray matter and of touch, etc., to the posterior white column cannot be accepted as demonstrated experimentally.
From the foregoing we may draw the following conclusions: -
1. Voluntary motor impulses from the cortex of the brain travel directly to the pyramidal tracts, and thence to the cells of the anterior gray columns. The fibres decussate in the medulla or in the upper part of the cord.
2. Motor impulses travel from the upper to the lower segments of the cord in the white fibres around the anterior gray columns.

Fig. 248. Diagram illustrating the course taken by the fibres in the spinal cord. (After Fick).
A, B and C represent oblique transverse sections of the cord, the tissue between being supposed to be transparent. At the lowest section (C), sensory nerve fibres (a) enter by the posterior root, and are connected with ganglion cells of the gray matter, and, through the posterior white column, with the brain (b).
Impulses arriving by the same posterior root may, to reach other parts, traverse the finer fibrils of the gray matter - shown by the fine lines.
When an impulse comes directly from the brain (voluntary centres) it adopts the direct routes (c or e), passing through the pyramidal tracts, to excite the motor ganglion cells of the cord to coordinated activity.
From many parts of the gray matter ganglion cells despatch impulses by the motor root (d).
Some white fibres only communicate between the cells of the various segments of the gray matter (f).
3. Various afferent impulses cross at once on entering the cord to the posterior gray columns of the other side, and then ascend by the neighboring white fibres of the posterior root zones, the direct cerebellar tract, the posterior median tract of Goll, and probably also by some of the white channels of the lateral column.
4. Besides their numerous thin protoplasmic connections in the various segments of the gray matter, all the cells of the cord are in communication with their more distant neighbors by means of the white fibres of the root zones.
Continue to:
My Books

