Immunity. Acquired Immunity
Description
This section is from the book "A Manual Of Pathology", by Guthrie McConnell. Also available from Amazon: A Manual Of Pathology.
Immunity. Acquired Immunity
By immunity is meant the power to resist invasion by microorganisms with the subsequent development of disease. An individual may be exposed to infection, but on account of some ability present may be able to resist and not acquire the disease.
The lack of resisting power is known as susceptibility.
One form of immunity is the natural, in which there is an inherited resisting power that is common to certain races of men or of lower living beings. The second type is the acquired, in which the immunity has been obtained after birth, and may be either active or passive.
Immunity is termed active when it results from the action of the cells within the invaded body, either in destroying the bacteria or in neutralizing their injurious products. It is that form which follows an attack of an infectious disease and which lasts for a varying period. It may be very brief, as in cholera; for a longer time, as in typhoid; or sometimes for life, as in small-pox. It may be due to accidental infection; to the inoculation of a weakened virus, as in vaccination; to the introduction into the body of bacterial products without the microorganism; or it may result from the inoculation of dead bacteria, as in bacterination.
Passive immunity is always acquired, never natural. It is that which is obtained by the introduction of the serum of an immunized animal into the body of a non-immune individual. The serum should always be introduced parenterally, that is, into the blood, subcutaneous tissues, or endothelial cavities, never by mouth, as it would then be acted upon by the digestive juices. It is supposed that in the serum of the first there is a substance that neutralizes the bacterial products in the blood of the infected animal.
It must be remembered that immunity is a comparatively relative term. An animal may resist an ordinary dose, yet succumb if a very large amount, either of bacteria or toxin, be administered. The degree of immunity may be reduced by unhygienic surroundings, by fatigue, by exposure to abnormal temperatures, abnormalities of diet, drugs, pre-existing disease, and by injuries.
Theories Of Acquired Immunity
Theories Of Acquired Immunity believed that the bacteria growing in the body used up some material that was necessary for their growth, and after dying left an unsuitable soil. This theory is not borne out by the facts.
2. Wernich And Chaveau
Wernich And Chaveau held that in the growth of bacteria 12 there were elaborated substances that inhibited their future development or activity. This theory also does not hold.

Fig. 68. - Leukocyte with Incorporated Bacilli, Illustratins Phagocytosis (Metchnikoff).
3. The Theory Of Phagocytosis
This is one of the most important, and is strongly supported by many well-known investigators. It, however, does not seem to be as satisfactory in general application as Ehrlich's "side-chain theory," which will be discussed later.
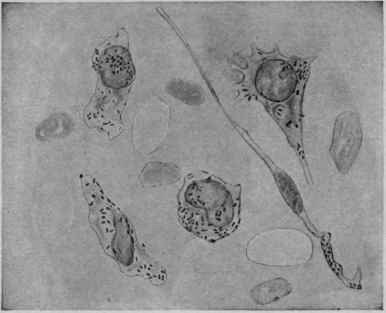
Fig. 69. - Phagocytosis: the Omentum Immediately after Injection of Typhoid Bacilli into a Rabbit.
Meshwork showing a macrophage, intermediate forms and a trailer, all containing intact bacilli (Buxton and Torrey).
There are certain cells in the body that have the power of ameboid motion, by means of which they are able to surround and take up bacteria and destroy them. These cells are known as phagocytes. Metchnikoff believes that immunity is the result of the positive and negative chemotaxis (power of attraction) existing between phagocytic cells and micro-organisms. He divides such cells into two classes:

Figs. 70 and 71. - Cells with Various Receptors or Haptophorous Groups of the First Order.
(a) Adapted to combination with the haptophorous groups (b) of various chemical compounds brought to them. It will be noted that there is no mechanism by which the toxophorous elements of the molecules (c) can be brought to the cell.
(1) Microphages - represented practically exclusively by the neutrophilic polymorphonuclear leukocytes.

Figs. 72 and 73. - Show the Regeneration of the Cell-hapto-phores or Receptors to Compensate for the Loss of Those Thrown Out of Service.
(2) Macrophages - large mononuclear leukocytes, endothelial cells lining serous membranes, and fixed mononuclear cells of the spleen follicles and lymph sinuses.
The most active are the microphages, as they have the power of independent motion. When the bacteria gain entrance into the body the phagocytes are attached, and they attempt to ingest and destroy the invaders. If the immunity of the animal is marked, many of the organisms will be found within the cells; if the immunity is slight, few cells will contain bacteria. At one time it was claimed that phagocytes could take up only dead bacteria, but it has been demonstrated that the leukocytes can take up living organisms. As a rule, the bacteria are ultimately destroyed, but on the other hand, the phagocyte may be the one to perish, and in this way permit a wide distribution of the invaders. Experiments have shown that the bacteria must be acted upon by a substance in the blood known as an opsonin, before the phagocyte is able to digest them.
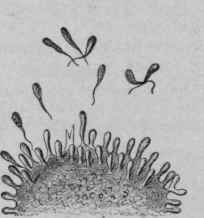
Fig. 74. - Shows the Number of Haptophores Regenerated by the Cell Becoming Excessive, They are Thrown Off into the Tissue Juice.

Fig. 75. - Explains What Antitoxins Are And How They Are Formed.
The liberated receptors in the tissue juice and in the blood possess identical combining affinities with those upon the cell, and meeting the adapted hapto-phorous elements in the blood, combine with them, thus keeping them from the cells.
4. Ehrlich's Lateral Chain Theory
This receives its name from its analogy to the benzole ring in chemistry with the accompanying lateral chains. For convenience terms are used that can be applied to formed bodies, although they cannot be demonstrated as such in the body juices.
In this theory it is claimed that immunity depends upon the presence or absence of "receptors," the equivalent of the chemical lateral chains. The normal or fixed receptor is that body attached to the cell by means of which the cell is acted upon by various substances, nutritive or otherwise. Each receptor is supposed to be so formed as to unite with a certain body of a definite character. When the action of the bacteria upon the tissue cells is considered, it is supposed that the poisonous products consist of two portions, the "haptophorous" and the "toxophorous" groups. The combination is thought to take place as follows: The haptophoric group unites with a certain definite receptor, and by so doing interferes with the normal function of the cell. At the same time the toxophoric group is able then to act directly upon the cell. If this group is very powerful the cell is destroyed, and if a sufficient number are involved the individual may die. When such a union occurs the receptors are of no further use. It has been found in such cases that in order to get back to the normal the cell will be so stimulated that new receptors similar to the' ones destroyed will be formed. As a rule, a great excess of receptors will result, and many of these will be cast forth into the circulation, becoming free receptors. These cast-off receptors are what constitute an antitoxin, and, on coming in contact with the toxin molecule, unite with the haptophorous portion and consequently render it harmless. It being attached to a free receptor, it is no longer able to combine with a fixed receptor.
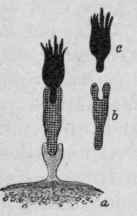
Fig. 76. - Combination of Cell (a), Amboceptor (b), and Complement (c).
The amboceptor may unite with the cell, but cannot affect it alone. The complement cannot unite with the cell except through the amboceptor, having no adaptation to the cell directly.
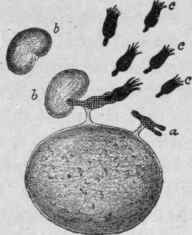
Fig. 77.
Cell with receptors of the second order (a) by which the cells fix useful molecules, of albumins, etc., on one hand (b), and zymogen molecules (c) on the other hand, and make use of the one substance through the action of the other.
In bacteriolysis and hemolysis, or cytolysis in general, conditions in which the destruction of actual cells is concerned, the destruction is brought about in a more complicated manner. Two other bodies than the cells are involved. One of these, known as the complement, is normally present in all serum. It is destroyed by a temperature of 550 C. for one-half hour, and is termed, therefore, thermolabile. The other, the immune body, or amboceptor, occurs in serum only as the result of the injection into the individual of the definite antigen. In other words, the amboceptor is specific in that it can combine only with that substance which gave rise to its formation. As it can resist heat up to 8o° C, it is termed thermostabile. In order that the cell be destroyed the complement unites with the specific amboceptor, which in turn joins with the fixed receptor, and the destructive action then occurs.
In active acquired immunity against foreign cells the invaded animal forms large amounts of amboceptor, which, being free within the blood, unite with and destroy the invading cells. If the supply is sufficient, the individual will recover. In passive acquired immunity large numbers of amboceptors in the serum from the immunized animal are directly introduced into the patient, and in this way effect a cure.
Continue to:
My Books

