Chapter 6, Note 8. Fixation Of Halogens On Conjugated Double Bonds
Description
This section is from the book "Research In Physiopathology As Basis Of Guided Chemotherapy With Special Application To Cancer", by Emanuel Revici. Also available from amazon: Research In Physiopathology
Chapter 6, Note 8. Fixation Of Halogens On Conjugated Double Bonds
The fact that the fixation of halogens at the conjugated double bonds occurs in two steps would explain the relative difficulty in a reverse reaction. As seen for butanediene, the halogen ions are first attached to the external carbons of the conjugated formation with the appearance of a double bond between the central carbons. It is only in a second step that two other halogens are bound to these central carbons too, thus completing the bond of halogens to all the carbons of the conjugated formation. (Fig. 248)
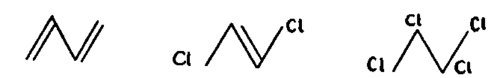
Fig. 248. Fixation of halogens at the conjugated double bonds takes place in two steps, with a displacement of the double bond in an intermediary position in the first phase, a fact which explains the nonreversibility of the process.
Chapter 6, Note 8a. Solvent Fractionation Of The Lipidic Constituents
In studying the biological role of the lipids, we recognized the importance of the forms under which the different lipids are present in the body, forms which seem to determine greatly their activity. A lipid changes its reactivity when it passes from the free form to one bound to other constituents. A first step in this study was the analytical separation of these forms. We distinguished thus four fundamental forms for acid lipids as well as for unsaponifiable fractions:
I. Free lipids, or those bound in such a labile physical form as to be able to take part directly in different reactions through their polar groups.
II. Lipids bound in cenapse with other constituents, that is, in a relatively labile form.
III. Lipids in combinations through their polar groups as esters or fats. This form represents usually a reserve or an inactive circulating form which can become active through hydrolysis.
IV. Lipids bound so firmly to other constituents as to be inseparable through solvents and to need saponification of the material in order to be liberated.
The first form would represent the functional form, the second a rapid available functional reserve, and the third, a reserve. The last would represent a stable combined form as part of the building of entities themselves.
We have utilized the differences in solubility of these various forms of lipids in order to separate them from the material to be studied, and thus to study their intervention in different normal and abnormal conditions. It must be emphasized that this separation concerns only the form under which the lipids are present in the organism and not their chemical constitution.
In spite of only a relative degree of accuracy in some separations, the differences noted from one sample to another are so manifest and so consistent that this method can be considered as an interesting and reliable source of information. We have, therefore, used this technique of separation for thousands of samples through the years.
According to the technique we devised, the material to be analyzed tissues, organs, entire organisms or only biological productsis finely divided in a blender. It is then extracted several times with ether under stirring, or in a Soxhlet apparatus. Under these conditions, ether removes the lipids present in the form of free lipids and neutral fats. This represents a mixture of the fractions I+ II. The residue is again extracted, this time with a mixture of 10% ethyl alcohol in ether which breaks the cenapses and separates the lipids previously bound in cenapse. The result represents fraction III. The residue is saponified with 10% KOH and extracted with ether. This represents fraction IV US or the unsaponifiable lipids of fraction IV. After acidification with tartaric acid, a new extraction is made which represents fraction IV LA or the acid lipids of this fraction.
The ether of fraction I + II is distilled and the residue treated with 85-90% alcohol concentration which dissolves selectively the free lipids as fraction I, and leaves the part formed by neutral fats as fraction II.
Fractions I, II and III are then saponified separately. The unsaponifiable fractions are extracted with ether, giving respectively the fractions I US, II US and III US. After acidification with tartaric acid, other extractions with ether are made which respectively represent fractions I LA, II LA and III LA. Each of these fractions is washed with distilled water, dried with anhydrous sodium sulfate and, after the ether is distilled off, the fractions are weighed.
For each material, we obtained thus four different fractions for the unsaponifiable part which we called the US fractions, and four for the saponifiable called the LA fractions. They correspond respectively to: I, free lipids; II, lipids present as esters or fats; III, lipids bound in cenapse; IV, lipids fixed in combinations which are liberated only through saponification. In the following examples, we chose different materials to illustrate the kind of information obtained through this method. Fig. 249 represents the results obtained in normal rats, in rats under abnormal conditions, as well as in tumors, all expressed as the 8 lipidic fractions.
In order to facilitate the comparison between normal and abnormal conditions, we chose for this example, rats between 180 and 200 gms. of weight, all males with one exception (case b). The data obtained were thus compared with that obtained for case (a) which corresponds to a normal male rat killed by ether. The analysis for the lipoacid fractions of this case shows a fraction (IV) at 3.5 mg./per gram of animal, the fraction III in cenapse at .2 mg./per gram, II corresponding to fats at 1.2 mg./per gram and the fraction I as free lipids at 8.1 mg./per gram of animal. For the unsaponifiable fraction, the fixed part IV is at 0.8 mg./per gram of animal, the III in cenapse at 2 mg., the II as fats at 0.8 mg., and the free part I at 7 mg./gram of animal.
The female rat (b) shows in general, lower values for the acid fractions and higher for each fraction of the unsaponifiable part. It is interesting to note that the fixed part (IV) in acid lipids is lower than in any male, a fact confirmed by other analyses. The increase in unsaponifiable part concerns especially the cenapse, fats and free lipid fractions. These differences are in agreement with the data concerning the relationship between lipids in males and females, as discussed in Chapter 6.
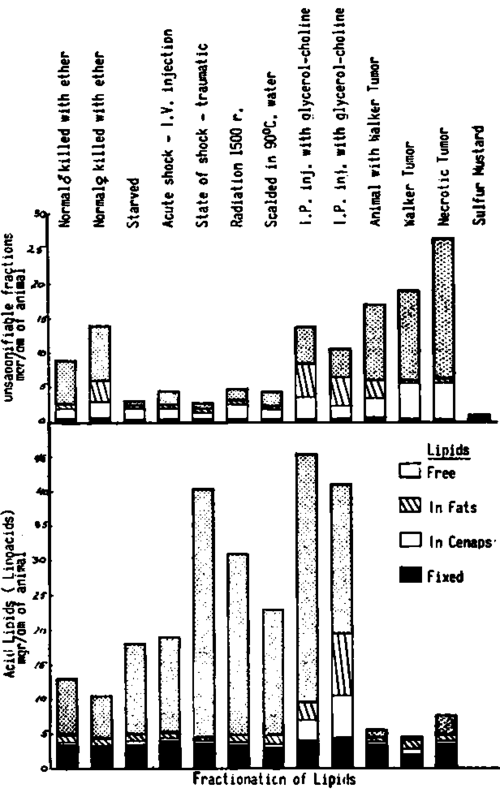
Fig. 249. Solvent separation between different lipoacids and unsaponifiable fractions in various animals and tumors. The values expressed in mgr/gr of animal show big variations concerning especially the free lipids. (Top fraction).
It is interesting to note the changes in starved animals as shown in case (c) where a marked decrease in the unsaponifiable fractions is seen, but with an increase however, in the acid part, especially in the free fraction (which reaches values of 13 mg./l gm.). Similar changes are seen in acute shock (d) induced by the intravenous injection of a rich culture of Esch. coli in broth. Although death occurred in less than 40 minutes, a marked increase in free fatty acids is seen with a corresponding decrease in the unsaponifiable fractions. The deviation from normal in the same direction appears still more manifest for the animal in a traumatic state of shock (induced by 700 falls in the Collip Noble drum), (e). The dying animal showed a marked increase in free fatty acid (37mg./l gm.) with a notable reduction in the unsaponifiable fractions. Cases with over 70 mg./l gm. of free fatty acid were also found. A lethal dose of radiation (1500r) (f), and caloric burns (g), were seen to induce similar changes.
Also of interest was the influence exerted by the intraperitoneal injection of glycerol and choline, with an impressing increase in all the fractions of the acid and unsaponifiable lipids, and of those in cenapse and fat fractions in (h). We chose example (i) of another animal submitted to the same treatment to show the extent of the concordance of the information furnished by this method.
It can be seen that almost under all conditions, the amount of fraction IV as obtained through saponification, changes very little. The amount present in cenapse and as neutral fats show more variations. Varying considerably from one case to the other, are the free lipids as fraction I which thus could be connected more directly with a pathogenic intervention of the lipids.
Another conclusion can be drawn from the analysis of these results. The high amount of free lipids often obtained under abnormal conditions cannot be considered to result only from a liberation of these lipids from the pre existing reserve as neutral fats or from the more labile cenapse form, since the total amount of the free lipids found is much greater than the sum of these forms. An appearance of new lipids, through synthesis, has become evident in these conditions.
Example (j) is the analysis of an animal with a 12-day old Walker tumor. Comparison with the normal controls shows a reduction of the acid lipidic part, with an increase in all the four fractions. The study of the tumor itself (k) shows still greater differences. The fixed fraction is smaller than in the total body of the animals, while the cenapse part is greatly increased. The free lipoacidic fraction is almost nil. For the unsaponifiable fractions, that in cenapse, and especially that corresponding to free lipids, is greatly increased. In the necrotic tumor all four lipoacid and unsaponifiable fractions are increased. The latter however, to a greater degree. Though the cenapse amount is high, an increase is most evident in the free fraction. Case (m) of an animal treated with sulfur mustard applied on the skin, appears particularly interesting. The animal, dying the 14th day, has almost no unsaponifiable fractions left (less than 1 mg./l gm.).
We used this method extensively throughout the years, in spite of its one limitationthe imperfect separation of neutral fats from free lipids.
The great concordance for all variations however, has practically overruled any objection of a failure of the separation of these free lipidic fractions.
Continue to:
My Books

