Steroid-Induced Growth In Hypophysectomized Rats
Description
This section is from the book "The Scientific Contributions Of The Ben May Laboratory For Cancer Research", by The University of Chicago. Also available from Amazon: The Scientific Contributions Of The Ben May Laboratory For Cancer Research.
Steroid-Induced Growth In Hypophysectomized Rats
These experiments were undertaken to correlate the growth-promoting activity of steroids with their molecular structure as an approach to the elucidation of mechanism of action of steroids. For the precise measurement of the effects of growth-promoters, it is required that growth other than that induced by the agent under assay shall be reduced to a minimum. This axiom had been largely neglected in earlier studies. Extraneous growth-promoters can come from within the animal (thyroid, pituitary) or from without (in the diet). Hypophysectomized rats maintained on a synthetic diet were found to fulfil these simple but important conditions. The hypophysectomized female rat is in a basal endocrine state; its genital tract is profoundly atrophic but retains great responsiveness (57) to growth-promoting steroids. The salient advantage over ovariectomized rats lies in the absence of the pituitary growth hormone.
Another advantage of this assay system is the possibility of testing androgenic and estrogenic effects in one animal. In previous studies, animals of two species (often birds and rodents were used) were necessary for full characterization of physiologic growth effects.
Steroids were administered to highly uniform hypophysectomized female rats maintained under strict climatic and dietary control. The principal test objects were (a) uterus; (b) vagina; (c) the giant sebaceous glands of the rat; (d) female prostate. Both the mass and the chemical types of growth (mucoprotein, keratin, etc.) were investigated.
Three structural factors in the steroid molecule are of high importance in determining physiologic activity: (a) the geometry of the molecule, (b) the number and site of functional groups, and (c) their state of oxidation.
In view of the importance of spatial considerations in determining activity of C19 steroids, it is reasonable to assume that close approximation to a specific surface is involved in exciting physiologic activity. If this be the case, the fit is ruined, for example, by a hydroxyl group on the back of the molecule at C-11, or on the front of the molecule at C-6. With respect to the ring structure, the extent and site of dehydrogenation are critical. Double bonds in ring A increase efficiency in promoting growth, and these increases are in direct relationship to the number of double bonds introduced. In ring B the introduction of a double bond at C-5 increases the effectiveness of a steroid as compared with the corresponding saturated compound, but a similar structure at C-6 abolishes activity.
Functional groups at C-3 and C-17 are of importance in the promotion of growth by steroids in the androstane series. As a generalization, hydro-genation of these oxygen functions or dehydrogenation at critical areas of the ring structure increase the quantitative efficacy of steroids in promoting growth. The position of double bonds and the state of oxidation at both C-3 and C-17 determine the qualitative type of growth which is reflected inter alia in cellular patterns which a compound in the androstane series induces in the vaginal epithelium. Thus 5-androstene-3β,17β-diol causes keratinization while 4-androstene-3β,17β-diol induces mucopro-tein synthesis in the vagina: the site of the double bond determines the metabolic pathway in the cell.
In order to evaluate the importance of the individual oxygen functions of steroid hormones in accelerating cell growth, a series of monofunctional steroids was synthesized and assayed (54). These monofunctional steroids possessed an oxygen function at position 3 or 17 but not at both sites (Fig. 8). The site of the functional group proved to be critical for the induction of growth by monofunctional steroids in the androstane series. Growth of the genital tract occurred in hypophysectomized rats only when the functional group was present at position 17; steroids with an identical group at position 3 were devoid of the capacity to promote growth. Further, the state of oxidation of the chemical group at C-17 was critical in promoting the growth process; whereas the presence of a β-oriented hydroxyl group at position 17 was effective in this regard, compounds with a ketone group at C-17 or C-3 were devoid of activity.
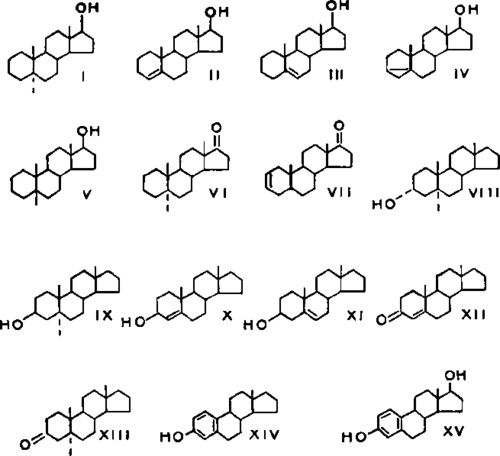
Fig. 8.
There remained some uncertainty whether the monofunctional steroid, androstan-17β-ol is biologically active per se or whether an oxygen function is introduced at position 3 in vivo. This difficulty was resolved (163) in an experiment wherein it was found that androstan-17β-ol and 3β-fluoroandrostan-17β-ol were highly active and equally effective in promoting growth of uterus, prostate, and of preputial glands. The fluorine atom at C-3 is tighdy bound and precludes oxygenation at this position. This result not only provides additional evidence for the significant role of the 17β-hydroxyl group in growth-promoting activity of compounds in the androstane series, but it indicates that fluorine can replace a hydrogen atom or hydrogen function at C-3 without appreciable loss of biological activity.
In sum, the capacity of the monofunctional steroids in the C19 series to induce growth of the uterus, vagina, and prostate of hypophysectomized female rats would seem to require (a) the presence of a hydroxyl group at (b) a specific site in a molecule which has (c) closely restricted geometric specifications. Hydrogen atoms at position 17 are of critical importance in steroid-induced growth by compounds in the androstane series. If a phenolic A-ring is present in monofunctional steroids, the 17β-hydroxyl group is not obligatory for growth. Proliferation of the uterus and vagina followed the administration of 17-deoxyestradiol.
The Impeded Estrogens
Hypophysectomized rats fed a synthetic ration free from growth-promoting steroids were employed in a study of a series of related phenolic estrogens (55) in the C18 series.
Two small groups of steroids in the estrane series-here designated as impeded estrogens-represent a class of compounds which differ from the majority of estrogenic substances in exerting certain unusual influences on growth of the uterus. The induction of these effects previously was considered to be peculiar to estriol. The unusual growth properties common to impeded estrogens are twofold: (i) After the threshold dosage required to initiate growth has been reached, the slope of the curve of increment of uterine weight in response to increased steroid dosage is very gradual rather than steep. (2) These compounds possess the ability to inhibit to a limited extent the uterine growth induced by estrone administered con-currendy.
The partial inhibition of estrone-induced growth of the uterus is confined to a critical dosage of the impeded estrogen and is overcome by increased dosage of the inhibitor which, of course, itself has estrogenic properties. Estrone-induced growth of the vagina is not inhibited by impeded estrogens, and testosterone does not lessen the amount of uterine growth evoked by the latter.
The impeded estrogens so far encountered are 3,17-dihydroxyestra-triene derivatives possessing either a ketone group at position 6 or a hydroxyl group (a or β) at position 16. Oxygenated functions at these positions in phenolic estrogens have special significance in the excitation and restraint of uterine growth unshared by similar groups at certain other sites of the estrane molecule.
Growth-Depressing Steroids
A study (56) was made of steroids related to progesterone, which depress the growth of specific tissues of the rat. As growth antagonists certain members of the pregnane series were found to have the following unique properties. The depression of growth is selective. Without interfering with body growth, some pregnanes cause a decrease in the size of adrenals and ovaries of intact young rats compared with the size of uninjected controls. In hypophysectomized rats these steroids inhibit the growth of the uterus when administered concurrendy with estrone or testosterone-hormones which are powerful stimulators of growth of the uterus.
Progesterone, 5-pregnene-3,20-dione, and deoxycorticosterone were found to be partial but significant inhibitors of estrone-induced growth of the uterus of hypophysectomized rats, but they did not depress the amount of growth elicited by testosterone.
The presence of both the 2-carbon side-chain at position 17 and an oxygenated function at position 3 is necessary for the inhibitory activity of progesterone. The introduction of a hydroxyl group in the progesterone molecule at position 6 (β), 11 (a), 16 (a), or 17 (a) destroyed its growth-inhibitory activity.
iiβ-Hydroxyprogesterone is weaker than progesterone in the inhibition of estrone-induced uterine growth; unlike progesterone, it has the capacity to depress growth elicited by testosterone.
The introduction of a fluorine atom in the 9 (a) position resulted in a striking increase of the growth-inhibitory activity of 11β-hydroxypro-gesterone and hydrocortisone.
9α-Fluoro-IIβ-hydroxyprogesterone is a partial but significant inhibitor of growth of the uterus in hypophysectomized rats injected simultaneously with large doses of estrone or testosterone. It is more powerful than progesterone in transforming the vaginal epithelium to mucous cells in the presence of estrone. Administered alone, the only detectable growth was a slight increase in size of the preputial glands.
9α-Fluoro-IIβ-hydroxyprogesterone depresses partially the growth of the adrenals, ovaries, and uterus in intact rats at a dose level which does not cause loss of body weight or inhibit body growth. Many rats treated with this compound in small amount had extensive mucification of the vagina resembling that of late pregnancy.
Continue to:
My Books

