Glandular Stroma. Part 5
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Glandular Stroma. Part 5
The myeloblasts were pipetted from the red cell layer and, in the most recent work, suspended in medium 199 (11) in a proportion of about 1 ml. cells in 10 ml. of suspension. With repeated centrifugation, the cells were washed in 3 successive changes of medium 199. On the basis of the packed-cell volume, noted in the graduated centrifuge tubes used in the washing process, the final cell suspension was parceled in lots depending on the requirements of the experiment. By repeated counts it was estimated that 1.0 ml. of cells, packed in the centrifugal field described, represented about 2.5 X 10^9 cells.
Cultures were established by introducing 0.05 ml. of packed cells into 5.0 ml. of culture fluid in 50 ml. Erlenmeyer flasks which were then closed with rubber stoppers. The cell concentration was thus approximately 2.5 X 10^7 cells per ml. of medium. The cultures were incubated at 37° C. on a rotary shaker (1) oscillating at 70 to 80 cycles per minute. Changes of culture fluid were made at intervals of 2 to 4 days. For this the cell suspensions were pipetted into 15 ml. graduated centrifuge tubes and spun at 150 X g for 10 minutes. The spent medium was removed for storage at -20° C, and the cells were resuspended in 5.0 ml. of fresh medium for further incubation as before.
Division of cultures was made when the number of cells was approximately 5 X 10^7 per ml. For this procedure, the cells of only 2.5 ml. of the culture were sedimented and then resuspended in 5.0 ml. of fresh medium. The remaining cells could be used for another culture or discarded. It was of considerable importance for continuity of the various characteristics of the cultures to maintain the concentration of cells at levels of about 2.5 X 10^7 to 5.0 X 10^7 per ml. The rate of cell proliferation was related to the conditions of culture, and the intervals between divisions varied accordingly.
Two measurements were made on the cultures at appropriate intervals: (1) enumeration of viable cells, and (2) determination of the number of myeloblastosis virus particles in the culture fluid. For the cell counts, aliquots of the well-mixed cells in culture were diluted with a preparation of trypan blue for identification of viable cells (1), and counts were made with a hemocytometer. Rapid enumeration of virus particles in small volumes of the culture fluids was made by a photometric procedure (12) for measurement of the adenosinetriphosphatase activity (18-15) associated with the virus particle. The standardization of the technique for accurate representation of virus particle number was controlled, as formerly (1, 2), by direct particle count with the electron microscope at suitable intervals. On the basis of this comparison of particle number with activity, the enzyme unit was chosen, as a matter of convenience, to correspond, on the average, to 9.15 X 10^8 particles per ml. of test preparation, of which a 50 X sample gave a reaction time of 1000 seconds.
The criteria of the processes occurring in the cultures were concerned, in this report, with the rate of cell growth and the rate of liberation of virus in terms of virus particles per cell per hour. The data describing cell growth represent the number of viable cells derived at a given time from 1.0 ml. of the initial culture. Ideally, these values would have been the sum of the respective values obtained by counts on all derivative cultures. It was not possible, however, to keep all the cells, and, in consequence, the actual data, obtained by counting the cells in a given culture, were adjusted by a calculation of the number of cells which would have been derived at the time of the count had all cultures been retained. This was accomplished by the multiplication of the number found in 1.0 ml. of the representative culture by the total number of cultures which may have existed had no cells been discarded. Virus liberation is expressed in an analogous way as the total virus derived from 1.0 ml. of the starting culture. The number of virus particles per ml. was measured in the spent culture fluids at each change of medium, and the values were accumulated from the preceding culture cycles. The resulting values were adjusted to account for the cells discarded as in the instance of the data on cell growth.
Results
The experiments presented here have been selected for exemplification of the major principles of cell growth and virus liberation observed in the study of myeloblasts during the past 3 years. Whereas wide variations encountered in the earlier experiments interfered with interpretation of the results, it is now possible by virtue of the findings to be described to perceive some elements of the trend of potentialities of the cells both for growth and for the synthesis and liberation of virus. The data are such that they are susceptible to mathematical interpretation, and analysis has been made of the data observed with the principal types of experiments.
Cell Growth and Virus Liberation in Cultures of Myeloblasts Fresh From the Chicken and Established in 50 Percent Chicken Serum in Medium 199 Supplemented With Added B Vitamins
The development of the culture medium currently in use has been guided by the results of numerous studies for various periods after preparation of cultures chiefly with cells fresh from the circulating blood. On the basis of the findings in work to be described in a later report, it was found that a critical requirement for the stabilization of cell growth was the presence in the medium of relatively high concentrations of folic acid.
The medium that has thus far yielded the best results consists of a mixture of equal parts of normal chicken serum and a modified form of medium 199. The modifications consisted in the increase in the concentration of (1) glucose from 1.0 to 5.0 gm. per liter; (2) the B vitamins (biotin, pantothenic acid, pyridoxal, thiamine, nicotinamide, and ribo-flavine) to the concentrations found in Eagle's (16) supplement; and (3) folic acid to 0.8 mg. per liter. Antibiotics were added to the whole medium in all experiments in concentrations of 50 units of penicillin and 50 µg. of streptomycin per ml. The serum was always heated for 30 minutes at 56° C. before preparation of the mixture.
The results of a typical experiment made with this medium are shown in text-figure 1. It is evident from inspection that cell proliferation conforms closely to a typical exponential growth rate function. This rate of growth was not attained immediately, but after a delay of approximately 4 days, after which the rate expression was linear throughout the remainder of the 30-day study.
Virus was first measured in the initial cultures after 3 days, and thereafter the agent was liberated at a rate apparently related to the number of cells present. Analysis of the data indicates that the rate of virus liberation, that is, the number of virus particles per cell in unit time, was constant in this experiment. This can be expressed by equation (1),
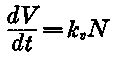
(1), in which V = the total number of virus particles at time, t, and N = the number of viable cells at time, t. The virus liberation constant (virus particles/cell/hour) is represented by kD. The equation describing the exponential rate of cell growth is
(2), where N0 = number of viable cells at the beginning of measurement; λ is the cell growth constant; and N = the number of cells at time, t. Substitution of N of equation (2) for N of equation (1), yields the relationship
(3).
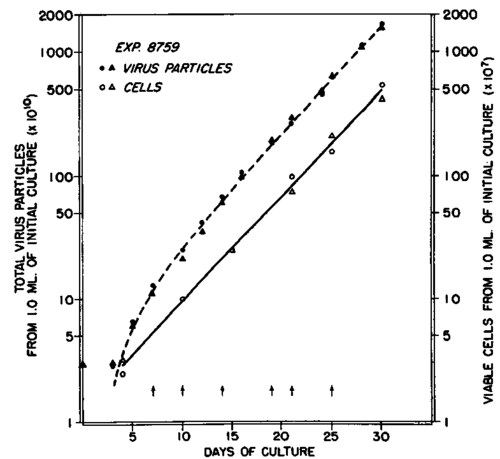
Text-figure 1. Cell growth and virus liberation in a culture of myeloblasts freshly obtained from the blood of a chicken with myeloblastosis. The culture fluid was 50 percent chicken serum in medium 199, with added glucose, B vitamins, and a concentration of 0.4 pg. of folic acid per ml. of whole medium. The continuous line was drawn by the method of least squares through the observed points representing cell growth. The dash line represents the expected total virus calculated by use of equation 4. Division of the cultures was made at the points indicated by the arrows.
On integration of equation (3), there is obtained the expression
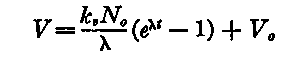
(4), in which V0 represents the number of virus particles present when t = 0.
When no virus is present at the beginning of the period of measurement, that is, when the cells have been washed free of the agent, V0 = 0. the expression applicable is
(5).
The applicability of these equations for interpretation of the results of the experiments with the myeloblasts can be tested by the fit of the observed data of experiment 8759 to the theoretical relationship. In text-figure 1 the continuous straight line was drawn by the method of least squares through the points representing the experimental data obtained by periodic counts of viable cells made during the period of exponential growth extending from 4 to 30 days. From the equation of this line the slope of the regression be,(be =log N\N0/days) was 0.0853. From this value, λ( λ= be/log e.24) was found to be 0.00819 hours-1. The number of cells, N0, at the beginning of exponential growth obtained from the intercept at 3 days, was 2.41 X 10^7 cells per ml. of culture.
Equation 4 is such that a plot of the experimentally determined number of virus particles, V, against the function (eλt - 1) should be linear. The results of this treatment of the data of experiment 8759 are shown in text-figure 2, in which the points represent the observed number of virus particles at various values of t. The line was drawn by the method of least squares. The slope of this line, kDN0/λ, was 81.06 X 10^9, and kD, was thus found to be 27.6 virus particles per cell per hour. V0, the intercept of the line, was 1.93 X 10^10. The various values are cited in table 1.
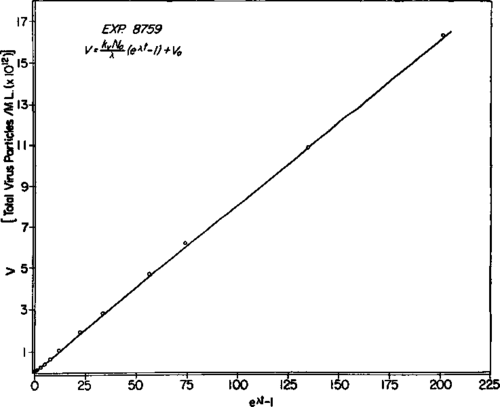
Text-figure 2. Total virus particles derived from 1 ml. of initial culture plotted according to equation 4. The open circles represent experimental values. The continuous line was drawn by the method of least squares, and the slope, kdN0/λ is 81.06 X 10^9. From this,kD, = 27.6 particles per cell per hour. The intercept, V0, of the line is at 19.3 X 10^9 particles.
The constants, X and k, determined in this way were used in equation 4 for calculations of the expected number of virus particles at different times, based on the original assumption that dV/dt = kvN, to yield the curve indicated by the dash line in text-figure 1. It is evident that the calculated curve is a very close fit to the experimental data. Tbus, it may be concluded that the rate of virus liberation was constant and was 27.6 virus particles per cell per hour. The generation time of the cells was 3.5 days (table 1).
Table 1. Cell growth and virus liberation constants of cultures of myeloblasts and a comparison with analogous data derived from results of studies on the Rous sarcoma by Prince (17)
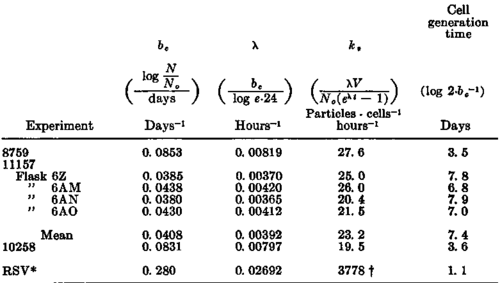
*RSV = calculations from the data of Prince given in figure S of (17). Pock-forming units per mg. dry weight per hour.
Continue to:
My Books

