Variation In Behavior Of Cells On Protracted Culture In Different Media
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Variation In Behavior Of Cells On Protracted Culture In Different Media
In the earliest experiments (1) the culture medium was 20 percent chicken serum and Gey's salt solution. The cells could be maintained for long periods in a state in which the rate of cell proliferation balanced approximately the rate of cell death. In this medium, virus was liberated first at a high rate followed by a gradual decline in the rate. In the first part of the curve of text-figure 3, it is seen that the rate of virus formation decreased from 28.6 to a level of 4.3 particles per cell per hour during the course of the study period of 148 days, while the cell population remained essentially constant. On the 148th day the cells were put into a mixture of equal parts of chicken serum and medium 199 without B vitamin supplement. Within a few days, an increase in cell population was noted, and on the 169th day the culture was divided. The course of cell proliferation thereafter is indicated by the increase in the number of viable cells. The fit of the experimental points to the straight line drawn through the observed points by the method of least squares showed that the rate of cell growth in the study period from 175 to 379 days was essentially exponential. This line represents a generation time of 7.7 days.
In contrast, the values representing virus liberation exhibited large and erratic variations. The nature and degree of these variations between 190 and 365 days can be seen from the relations derived by analyses similar to those of the preceding section and illustrated also in text-figure 4. The dash lines (t = 0 on day 190) represent the expected relations on the assumption of constant rates of 40, 20, 10, 5, and 1 virus particles liberated per cell per hour.
The results of text-figure 4 show that, whereas the medium supported an exponential rate of growth of the cells, the rate of virus synthesis and liberation, nevertheless, varied greatly. Although the exact basis for this variation was not discernible, it was realized that some or much of it was related to the use of different batches of serum during the study period (1). In such a system, however, it could not be predicted in advance that the variation might not be due to actual changes in the properties of the cells due to prolonged maintenance in culture. That this was probably not the case was shown by the results of the following study.
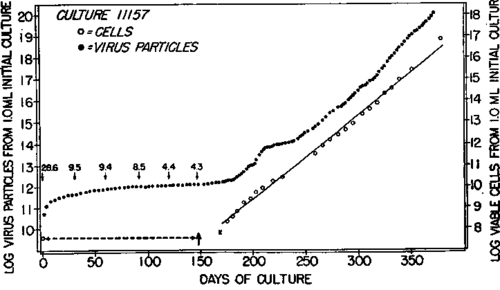
Text-figure 3. Behavior of a long-term culture of myeloblasts, first (1), in a medium of 20 percent chicken serum in Gey's salt solution for 148 days, and then, in a mixture of 50 percent chicken serum in medium 199 with added glucose but without added B vitamins. The dash line signifies lack of increase in cell population in the initial period. At 169 days (indicated by the cross) cell growth was sufficient to permit division of the culture, and the culture was divided 33 times thereafter. The open symbols represent the results derived from cell counts, and the continuous line was drawn by the method of least squares. The numbers above the virus data (closed circles) indicate rate of virus liberation per cell per hour at the intervals shown by the corresponding arrows from 0 to 148 days.
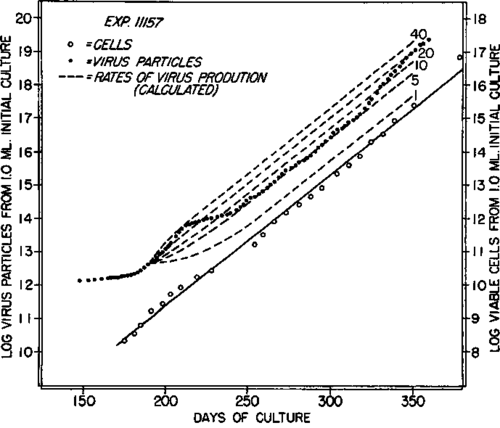
Text-figure 4. Analysis by application of equation 4 of the data of a portion of the experiment of text-figure 3. The dash lines were drawn from day 190 to represent the relations expected on the assumption of constant rates of virus liberation of 40, 20, 10, 5, and 1 particles per cell per hour.
Continue to:
My Books

