Growth Curve Of Virus In Suckling Hamsters
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Growth Curve Of Virus In Suckling Hamsters
Three experiments were carried out to determine the growth curve of polyoma virus after subcutaneous inoculation into suckling hamsters (11). Groups of suckling hamsters 2 to 4 days of age were inoculated with 0.1 ml. of undiluted infectious tissue-culture fluid given subcutaneously on the back. At prescribed intervals, groups of 3 to 6 animals, each from a separate Utter, were sacrified, the organs examined in the gross for the characteristic hemangiomatous and sarcomatous tumors, and various organs removed for testing. In some instances, individual organs were tested; in others, a pool was made of the heart, lungs, liver, and kidneys. Each suspension represented corresponding tissues of a number of animals. In experiment #H-1 the virus was assayed by titration in mouse embryo tissue cultures, and in the other two experiments, the titrations were done by the MAP procedure.
Table 9 presents the results of these 3 experiments, giving the course of viral growth, antibody response, and the time of appearance of grossly visible tumors. The inoculums used in the experiments were highly pathogenic; some animals demonstrated grossly visible tumors 10 to 14 days after inoculation, and all animals killed 21 days or more after inoculation demonstrated multiple tumors. The tests for infectious virus titer indicated that a peak of infectivity was reached within 3 to 6 days after inoculation, with highest titers in the kidneys, heart, and liver. After the peak of infectivity, virus titer declined rapidly. In experiment #H-2, the titer in most organs was very low by 7 to 10 days, and, in both experiments #H-1 and #H-2, the pooled organs were generally noninfectious after the end of the 2d week of infection.
Antibody response in the suckling hamsters was prompt and of high titer. HI antibody appeared within 7 to 10 days after inoculation, and neutralizing antibody was also present by the 10th day. Complement-fixing antibody appeared slightly later, at 10 to 14 days.
Additional Attempts To Recover Virus From Hamster Tumors
The data described in the preceding section indicated that tumors removed 3 weeks or later after inoculation were generally noninfectious. Since the previous tests were done with pools of tissues from several animals, additional tests were made with tumors from individual hamsters. Tumorous organs from hamsters sacrificed 16 to 82 days after inoculation were made to 10 percent suspensions in Eagle's medium and tested for infectivity by the MAP and/or mouse embryo tissue-culture procedures. The results are summarized in table 10. Three of the 4 tumors removed at 16 to 17 days contained detectable viral infectivity; however, the quantity was small and required, in one case, blind passage in tissue culture to produce cytopathic changes, and, in the other 2 cases, relatively prolonged observation periods. Of the tumors removed at 43 days, only 1 of the 5 contained detectable virus, and in this instance only in very low titer. The 5 tumors tested at 82 days were negative for virus in both tests, and the tumor extracts did not produce tumors or HI antibody when inoculated into suckling hamsters.
Serial Passage Of Polyoma Virus In Suckling Hamsters
The titers of virus obtained at the peak of virus activity in the growth curve experiments were not sufficiently high to establish unequivocably that viral multiplication had occurred in the hamsters. To establish this point, two serial passage lines of polyoma virus were carried out in suckling hamsters. Line 1 was initiated with the pooled-organ suspension taken at 7 days in experiment #H-1 (table 9), and passage line 2 was initiated with the 3-day pooled organs in experiment #H-2 (table 9). Serial passages were made at 5- to 7-day intervals by preparing suspensions of the heart, lungs, liver, and kidneys of 4 suckling hamsters that had been inoculated as 2- to 4-day-old animals, and the undiluted clarified suspension was inoculated into 2 litters of hamsters. The hamsters which were not sacrificed for passage were maintained 5 weeks for observation of tumor incidence. Many such hamsters were also bled at the time of sacrifice to determine if there was an antibody response to the inoculum. Each suspension was also inoculated into 5 weanling mice as a parallel test for viral infectivity. The 2 lines were successfully carried through 19 and 16 serial passages, respectively (table 11). Tumors were observed sporadically throughout the passage series, but were not observed after the virus had been lost. The low incidence of tumors indicated that the virus was not becoming highly adapted to bo oncogenic for tho hamster, at least in the dosages used in the form of hamster tissue suspensions. The suckling hamster suspension at the 11th passage level was titrated by the MAP procedure and yielded a titer of 10s-1 ID50 per 0.2 ml., which is comparable to those obtained in the growth curve experiments. This suspension was also inoculated into mouse embryo tissue culture, and virus was recovered; the tissue-culture fluid when inoculated undiluted into suckling hamsters was highly oncogenic, and produced tumors in each of the 11 surviving hamsters within a period of 35 days.
Table 9. Development Of Virus And Antibody After Subcutaneous Inoculation Of Suckling Hamsters With Polyoma Virus
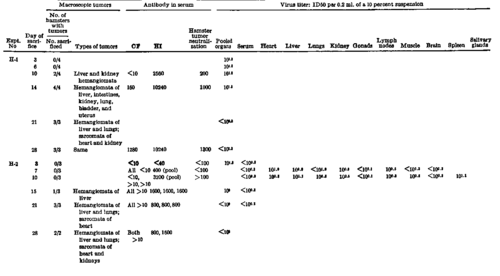

Table 10. Recovery Of Virus From Hamster Tumors
Results of virus assays on tumor extract | |||||||||
Type of tumor | Removal after in-Dculation {day) | MAP test * | Mouse embryo tissue culture | Suckling hamsters (85-day test) | |||||
10^0 | 10^-1 | 10^-2 | 10^-3 | 10^-4 | 10^0 | Tumors | HI antibody | ||
Hemangiomata of liver and lung | 16 | 2/2 | + (17-24 Days) | ||||||
Hemangiomata of liver and lung | 16 | 1/2 | + (17-21 Days) | ||||||
Hemangiomata of liver and lung | 16 | 0/2 | - (Blind passage) | ||||||
Hemangiomata of liver | 17 | + (Blind passage) | |||||||
Sarcoma of heart | 43 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | |||
Hemangiomata of liver | 43 | 2/3 | 0/3 | 0/3 | 0/3 | 0/3 | |||
Hemangiomata of liver, sarcoma of heart, and | 43 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | - (Blind passage) | ||
subcutaneous sarcoma | |||||||||
Sarcoma of heart | 43 | 0/5 | 0/3 | 0/3 | 0/3 | 0/3 | - (Blind passage) | ||
Hemangiomata of liver, sarcoma of heart | 43 | 0/6 | 0/3 | 0/3 | 0/3 | 0/3 | - (Blind passage) | ||
Hemangiomata of liver | 82 | 0/3 | - (Blind passage) | 0/13 | 0/13 | ||||
Sarcoma of heart | 82 | 0/3 | - (Blind passage) | 0/16 | 0/16 | ||||
Sarcoma of uterus | 82 | 0/3 | - (Blind passage) | 0/14 | 0/14 | ||||
Subcutaneous sarcoma | 82 | 0/3 | - (Blind passage) | 0/7 | 0/7 | ||||
Subcutaneous sarcoma | 82 | 0/3 | - (Blind passage) | 0/6 | 0/6 | ||||
*Numerator - number of mice developing antibody (35-day test). Denominator - number of mice Inoculated with dilution of tissue extract.
Numerator - number of hamsters developing macroscopic tumors. Denominator = number surviving 35 days after subcutaneous Inoculation with 0.1 ml. of 10 percent tumor extract. Numerator - number of hamsters developing antibody. Denominator » number surviving 35 days.
Table 11. Tumor Response And Tests For Virus In Serial Passage Lines Of Polyoma Virus In Suckling Hamsters
Passage line 1 Suckling hamsters MAP test | Passage line 2 Suckling hamsters MAP test | ||||||
Passage | Day passed | Tumors* | HI antibody | HI antibody | Tumors* | HI antibody | HI anti-body |
2-10 | 5-7 | 10/30 | 7/7 | 45/45 | 6/35 | 6/6 | 44/44 |
11-15 | 5 | 7/17 | 11/11 | 21/21 | 4/15 | 13/13 | 22/23 |
16 | 5 | 2/2 | 5/5 | 0/8 | 8/8 | 5/5 | |
17 | 5 | 0/8 | 7/8 | 5/5 | 0/1 | 0/1 | 0/5 |
18 | 5 | - | 4/5 | 0/1 | 0/1 | 0/5 | |
19 | 5 | 0/2 | 0/2 | 4/5 | 0/7 | 0/7 | 0/5 |
20 | 5 | 0/5 | 0/5 | 0/4 | 0/4 | 0/3 | 0/5 |
*Numerator = number developing gross tumors during 35-day observation period. Denominator - number observed.
Numerator - number of animals developing HI antibody (> 100 titer). Denominator - number tested.
Discussion
The results presented here indicate that the pattern of growth of polyoma virus in mice and hamsters follows that of the majority of viruses of acute illnesses, i.e., maximal growth of virus within a few days or weeks, followed by decline or disappearance of detectable virus. However, the fact that the peak of viral activity significantly precedes the appearance of the overt manifestation of the infection-the tumor-carries many important implications.
In an examination of the mechanism of tumorigenesis by viruses, two basic patterns must be considered. Either the virus infection induces a cellular alteration which is later expressed as tumorous growth independent of persistence of virus in the cells, or unrestrained cellular growth occurs as a reaction to continual activity of virus in the cells. The growth patterns of polyoma virus, particularly that in the hamster, are highly suggestive of the former mechanism and imply that searches for the cellular alteration leading to tumor formation should concentrate on the early period after infection.
It will be important to determine if differences in tumor response of mice under different experimental conditions can be related to differences in time, height, or duration of peak virus titers or to the titer of persistent virus. The experiments described here demonstrated marked differences in tumor response, but the titration data are not adequate to justify any conclusions and can only serve to suggest hypotheses for further testing. Of the mouse experiments reported here, only experiment 1 represented a highly oncogenic system and must form the basis for comparison. The contrast between this experiment and the growth curve in weanling mice was great, both in magnitude of peak virus growth and in titer of persistent virus. However, a correlation of growth pattern with the marked difference in tumor response between experiments 1 and 2, both conducted in newborn mice, is not as apparent; the most noteworthy difference between the growth curves in these two experiments was the titer of virus in the 2d through 5th months, which was lower in the experiment with poor tumor response. This observation, if confirmed, would support the concept that tumor induction is a result of continued presence of virus.
Continue to:
My Books

