Growth Curves of Polyoma Virus in Mice and Hamsters
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Growth Curves of Polyoma Virus in Mice and Hamsters
Knowledge of the pattern of growth of an infectious agent is important to an understanding of pathogenesis and natural history of the infection. This report presents data on the general patterns of growth, organ distribution, and persistence of polyoma virus when inoculated parenterally into newborn mice, weanling mice, and suckling hamsters.
Materials And Methods
Virus
Inoculations for all experiments were infected mouse embryo tissue-culture fluids; procedures for preparation of virus stocks have been described elsewhere (1). Unless otherwise stated, virus used was the LID-1 strain of polyoma virus (1).
Animals
Mice were of the general purpose randombred Swiss albino strain, obtained from the Animal Production Section of the National Institutes of Health; the mice in this colony have been shown to be free of polyoma virus infection as measured by absence of antibody in more than 1,700 adult mice of various strains tested (2, 3).
Pregnant Syrian hamsters were received from the Animal Production Section of the National Institutes of Health and from several commercial hamsteries. Serums from representative animals from each colony were tested for hemagglutination-inhibiting (HI) antibody to polyoma virus, and none were positive.
Preparation Of Materials For Virus Assay
Animals were exsanguinated by cardiac puncture under ether or chloroform anesthesia. Individual organs designated for separate testing were removed aseptically with separate instruments. Tissues were weighed,homogenizedinamortarorTen Broeck tissue grinder, and made into 10 percent suspensions in Eagle's basal medium (4). The suspensions were clarified by centrifugation at 2500 rpm for 15 minutes in a refrigerated centrifuge. Generally, tissues were assayed for virus on the day of harvest; otherwise they were stored at -60 or -20° C, usually as whole organs, until tested.
Urines and mouth swabs were collected antemortem. In the initial experiments, a single voided specimen of urine was diluted in 5 ml. of Eagle's medium; after the importance of urinary excretion had been found, precise dilutions of urine were obtained by collecting the urine in a weighed watch glass, reweighing on a scale with a sensitivity of 5 mg., and rinsing out the glass with an appropriate volume of Eagle's medium. Mouth swabs were taken by twisting a cotton-tipped applicator stick in the mouth; the swab was then squeezed out in a vial containing 5 ml. of Eagle's medium.
Infectivity Titrations
With the exception of one experiment, noted in the text, all assays for infectivity were made by the mouse antibody production (MAP) procedure (1). Serial tenfold dilutions of test material were made in Eagle's medium, and were inoculated intraperitoneally into groups of weanling mice of the general purpose stock in volumes of 0.2 ml. per mouse. In routine titrations, 3 mice were used per serial dilution. The mice were tested individually for development of HI antibody response 35 days after injection. Control mice, consisting of uninoculated and saline-injected mice, some of which were placed in cage contact with test mice, were incorporated into each MAP test. In the tests reported here, a total of 1,082 control mice were used, with serologic conversion in 10, which represented a false positive rate of 1 percent.
Titers of tissue-culture fluids and hamster tissue suspensions were calculated by the Reed-Muench method (6) and are expressed as the number of 50 percent infectious doses (ID50) per 0.2 ml. of 10 percent tissue suspension. In tests of mouse tissues, harvested after the development of high-titer antibody, titrations were erratic, as described in the text, and the Reed-Muench formula was not applicable. Consequently, titers in the mouse experiments are given as the highest dilution of test material which induced antibody response in more than half the recipient animals.
Hemagglutination (Ha) Test
Tissue suspensions were tested for ability to hemagglutinate guinea-pig erythrocytes, with the use of standard procedures described previously (1, 6). In order to "unmask" hemagglutinin from nonspecific inhibitors in tissue extracts (7), the tissue suspensions were treated with receptor-destroying enzyme (RDE) prior to testing. This treatment was carried out by mixing equal volumes of tissue suspension and undiluted RDE preparation consisting of unpurified filtrate of Vibrio cholerae broth cultures; after incubation at 37° C. for 1 hour, the mixture was heated at 56° C. for 30 minutes, and diluted to a final concentration of 1 percent tissue with pH. 7.2 phosphate-buffered saline.
Serologic Tests
HI and complement-fixation (CF) tests were carried out, as described previously (2, 8), with antigens prepared with the LID-1 strain. Virus neutralization tests in suckling hamsters were carried out by the following procedure: Tissue-culture fluid infected with the LID-1 strain was diluted 1:5 and mixed with an equal volume of tenfold serum dilutions; the serums had been previously heated at 56° C. for 30 minutes. Virus controls consisted of virus mixed with saline or control mouse serum. The mixtures were held at room temperature for 30 minutes, and 0.2 ml. volumes of each test mixture were inoculated subcutaneously into 1 or 2 Utters of 2- to 4-day-old hamsters. Animals dying after the 14th day and all animals surviving at 20 days were autop-sied and examined macroscopically for tumors. Neutralization titers of serums were calculated by the Reed-Muench method, calculating the dilution of serum which would reduce tumor incidence by 50 percentage units; for example, since, in the neutralization test on the serums in the first experiment listed in table 9, the virus controls gave 74 percent tumor response (20/27 animals), the titers were calculated as the dilution of serum which would reduce tumor incidence to 24 percent.
Results
Growth of Virus After Inoculation of Newborn Mice With High Dosage
To determine the growth pattern of virus under the conditions usually employed to obtain tumor response-inoculation of newborn mice with high dosage of virus-two chief experiments were carried out. Mice less than 12 hours old were inoculated subcutaneously with 0.1 or 0.2 ml. of a 1:10 dilution of infectious mouse embryo tissue-culture fluid, representing approximately 10^6 mouse ID50 per mouse. The inoculum in experiment 1 was the fifth tissue-culture passage of the LID-1 strain, and in experiment 2 was the seventh passage of the same strain. The tumor response obtained was markedly different between the two experiments : By 150 days after inoculation, 48 of 54 surviving mice in experiment
I had developed parotid tumors, while none of 21 mice in experiment 2 developed grossly visible tumors by the 150th day, and of 9 mice held for
II months, only 2 developed tumors. The reason for this difference in response is not known.
Tables 1 and 2 present the infectivity and HA titers of selected tissues obtained at various intervals after inoculation. High infectivity titers were attained by the 7th to 9th day after inoculation, and tended to remain at comparable or slightly lower levels through the 20th to 30th day. Thereafter the titers were lower and more variable.
During the initial peak phase, virus was widely distributed throughout the viscera, and the titers were sufficiently higher than that expected by their content of blood to indicate that the virus was propagating in each organ tested. Highest titers were observed in the kidneys, salivary glands, liver, lungs, and spleen, with somewhat lower titers in thymus, brain, and blood.
After the 2d month of infection, there was a marked difference between the two experiments with respect to the titer of virus detected. The mice in experiment 1 regularly contained virus in the salivary glands, with titers of 10^2 to 10^4, while the mice in experiment 2 were less often positive at the level of testing, and the titers in the positive mice were very low.
Table 1. Growth Of Polyoma Virus In Various Organs After Inoculation Into Newborn Swiss Mice: Experiment 1
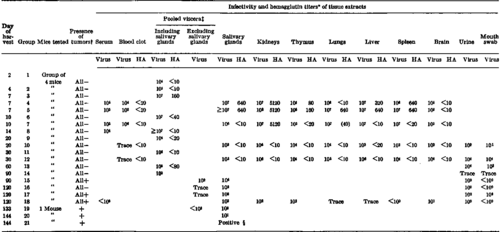
*Infectivity and HA titers of serum, blood clot, and tissues are expressed per 0.2 ml. of 10 percent suspension. Infectivity titers of urine and mouth swab are expressed per 0.2 ml. of a 5 ml. volume of saline solution in which was collected a single voided specimen of urine or in which a mouth swab was rinsed. HA titers determined on suspensions after treatment with RDE.
Presence of tumors as determined by gross examination.
Pool of kidneys, adrenals, thymus, lungs, liver, spleen, pancreas, mesenteric node, gonads, and brain. §Titer could not be evaluated. See table 5.
Table 2. -Growth Of Polyoma Virus In Various Organs After Inoculation Into Newborn Swiss Mice: Experiment 2

*Infectivity and HA titers of tissue suspensions expressed per 0.2 ml. of 10 percent suspension. Infectivity titers of urine expressed per 0.2 ml. of a 6 ml. volume of saline solution in which was collected a single voided specimen of urine. HA titers determined on suspensions after treatment with RDE.
Pool of parotid and submaxillary salivary glands, kidneys, adrenals, thymus, lungs, liver, spleen, pancreas, mesenteric node, gonads, and brain. Presence of tumors as determined by gross examination. §Pooled serums of mice in the group.
As expected from the high titers of virus in salivary glands and kidneys, the mouth swabs and urine specimens were frequently infectious and occasionally in high titer. More extensive data on excretion of virus have been presented elsewhere (3).
Table 3 presents results of a similar experiment, which was carried out to compare the early phase of growth in an inbred mouse strain with that previously described in Swiss mice. Pregnant DBA/2Lw3 mice were tested for HI antibody, and the litters of antibody-negative mice were inoculated when less than 12 hours old. The inoculum was 108-7 mouse ID50 of fifth mouse embryo tissue-culture passage of the LID-1 strain, given subcutaneously. The tissue suspensions were assayed by HA tests. The results were comparable to those in experiments 1 and 2, with high HA titers on the 8th day, and maximal HA titers in the kidneys. It is noteworthy that 4 of the urine samples gave HA at a 1:100 dilution. While these specimens were not tested for specificity of the HA by HI tests, these reactions probably represented specific viral HA, since tests of normal urine have not given positive results at lower dilutions, and in other experiments high titers of hemagglutinin have been found in infected mouse urine, with specific inhibition by polyoma antiserum.
Table 3. Hemagglutinin Titers In Tissues Of Dba Mice Inoculated With Polyoma Virus As Newborns
Time after inoculation (.days) | Hemagglutinin titer* | ||||||
Mice tested | Pooled organs | Salivary glands | Kidney Thymus | Liver | Urine | ||
5 | Pool of 3 | 200 | <100 | ||||
8 | Individual | <100 | 5120 | <100 | 400 | <100 | |
8 | Individual | 400 | 5120 | <100 | 100 | ||
8 | Individual | 400 | 2560 | 100 | <100 | 100 | |
12 | Individual | <100 | <100 | <100 | <100 | 100 | |
14 | Individual | <100 | <100 | <100 | <100 | 100 | |
*Titer per 0.2 ml. of 10 percent tissue suspension, and per 0.2 ml. of urine. All titers determined after RDE treatment.
Continue to:
My Books

