Maturation And Release Of The Tumor Cell Virus. Continued
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Maturation And Release Of The Tumor Cell Virus. Continued
Summary And Conclusions
The significant points to be brought out in the present discussion are as follows:
1. In direct correlation with biologic evidence, the electron microscopic evidence shows that tumor virus particle formation, in precursory or elementary form, occurs in association with genie material in the nucleus, and with enzymatic sites in the mitochondria, resulting in modification of these organelles.
2. At least in several of the viruses studied in the present investigation, the elementary viral substance is similar in size and structure to the elementary substance of the normal genetic coded material, that is, the viral unit substance is composed of twisted filaments of the dimension of the alpha helix.
3. In the formation of the tumor virus, at least in the Gross leukemia agent, the elementary filamentous units enter into the formation of the coat as well as nucleoid of the virus. Hence the viral coded information exists in all parts of the mature virus.
4. It can now be explained why investigations to date have not shown entering or migrating virus particles, and perhaps why virus particles as such have not been visualized in human tumor cells.
5. As shown in several instances mentioned, and in one instance of filamentous material continuous from nucleoplasm through the pore channel and cytoplasm up to the point of virus particle formation at the surface, it is in the twisted filamentous form that the leukemia, and possibly polyoma, virus material exists in the nuclear and cytoplasmic matrices. Replication would appear to consist of formation of an excess of filamentous units. These in turn form aggregates or crystals within the cell at optimal sites under the usual laws of crystal formation and concentration phenomena. The mature virus is apparently formed only at the cell surface prior to release.
6. The question arises as to whether the modifications of the affected cell are brought about by changes in its own coded information due to propinquity of viral information, or whether the viral information controls the pathways and processes of the cell, bringing about local modifications directly related to its formation and release. The latter seems more logical. The entry and initial migration could be as for any substance, be it neutral or active, i.e., by pinocytosis and/or membrane flow. The replication and release apparently are brought about by changes in normal pathways due to the new information contained in the viral material. The cytopathic or noncytopathic effect of the virus would thus also depend upon the closeness of fit of viral and cellular information codes.
7. Now that it is possible to visualize the structure of the viral components at the molecular level, it becomes imperative to devise methods for distinguishing between normal and viral units in nucleus and cytoplasm, as well as within the mature virus itself. Included in the information to be thus derived would be that on latency, mutation, and contamination. A hopeful step in this direction has been provided by the chromium stain developed by Bullivant and Hotchin (27). In preliminary experiments there is some indication that it is possible to develop a method that specifically stains the viral material only.
Whether this particular method is successful or not, the results obtained so far indicate that the next step is not merely to look for or recognize large viral particles, but to distinguish viral from normal subunits at the level of the most elementary structures of the cellular organelles.
Plate 55
Figure 12. Portion of leukemia tumor cell .showing nucleus (nu), cytoplasm (cy), nuclear envelope with perinuclear cisterna (ci) interrupted by numerous pores (arrows). Unoriented fine filaments are seen in nucleoplasm (f1). In pore channel (U) and on cytoplasmic side of pore, filamentous material (f3) appears in parallel array. Occasionally strands of perinuclear filaments (f4) are seen to be continuous with the cortical cytoplasmic filamentous material (f5, f6). Virus particle (v) shown at cell border (pm). Note its relation to peripheral cytoplasmic filaments. X 120,000.
Figure 13. Leukemia tumor cell showing accumulated filamentous material (f, f1) at subplastnalemmal cytoplasmic region. Plasma membrane (pm) and fine filaments enter in virus particle formation, as in early stage (v1), and in more mature form (v2), where filamentous material (f2) is seen in direct relation with virus particle. X 140,000.
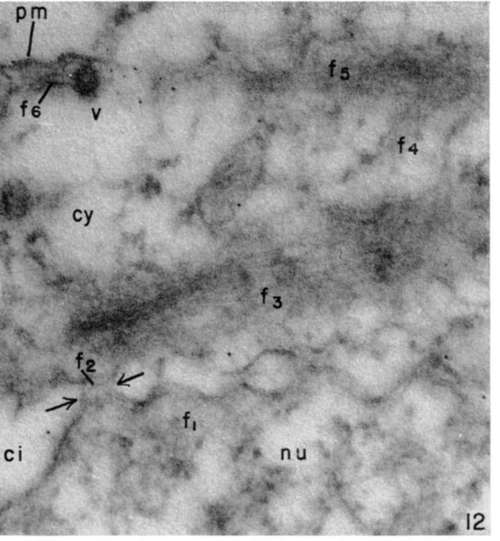
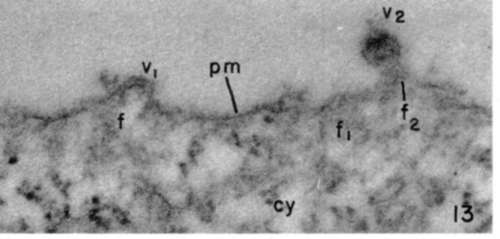
Plate 50
Figure 14. Region between two leukemia tumor cells. Four virus particles (v) are seen in interstitia, sometimes appearing to be connected with plasma membranes (pm) (arrow). "Inclusion body" (ib) shows a filamentous matrix with rows of tiny dense particles. Lipide clumps (1) and normal mitochondrion (mi) are also seen. X 120,000.
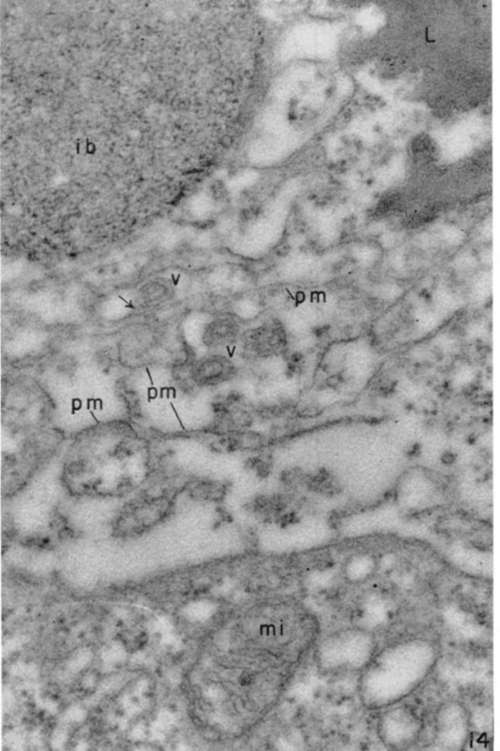
Plate 57
Figure 15. Edge of leukemia tumor cell showing virus panicle (v) attached by long connection to plasma membrane (pm). Fine filamentous material If) from cortical cytoplasm is seen to be continuous with virus attachment. X 100,000.
Figure 16. Periphery of leukemia tumor cell showing infoldings of plasma membrane (pm). partially formed virus particle (v1), and more advanced particle (v2) in which cross striatums can be seen in virus coal. X 80.000.
Figure 17. High magnificat ion of newly formed leukemia virus particle. Note complex membrane and spiral or coiled internal array of filamentous material which appears to connect with plasma membrane (pm). X 440,000.
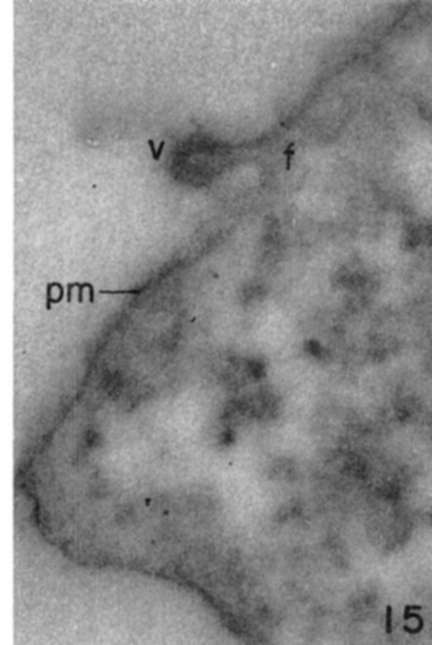
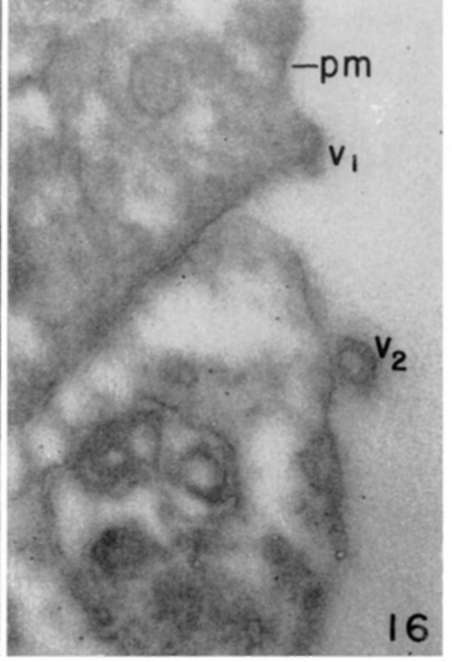
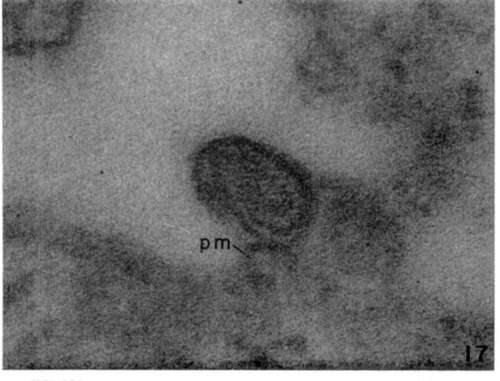
Plate 58
Figures 18 through 21 show mature and young leukemia-type viruses found at borders of cells of a polyoma parotid tumor (cf. 3).
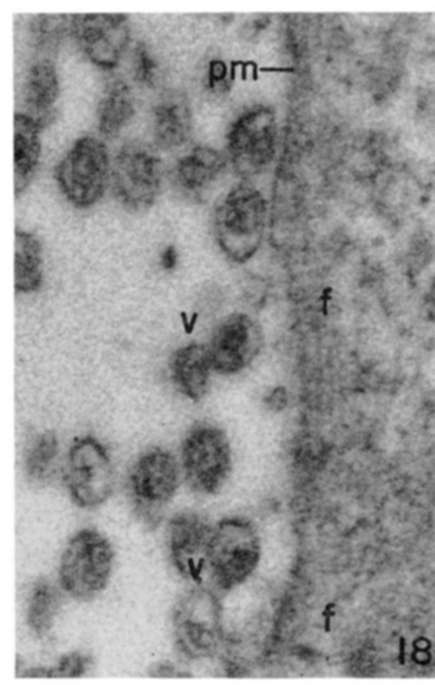
Figure 18. Note parallel arrays of fine filaments along edge of cell. Virus particles (v) are seen outside plasma membrane (pm) in intercellular space. X 120,000.
Figure 19. Similar to figure 18 but displaying a half-formed virus particle (vi) and its relation to plasma membrane (pm) and to fine filamentous material (f) in cortical cytoplasm. X 120,000.
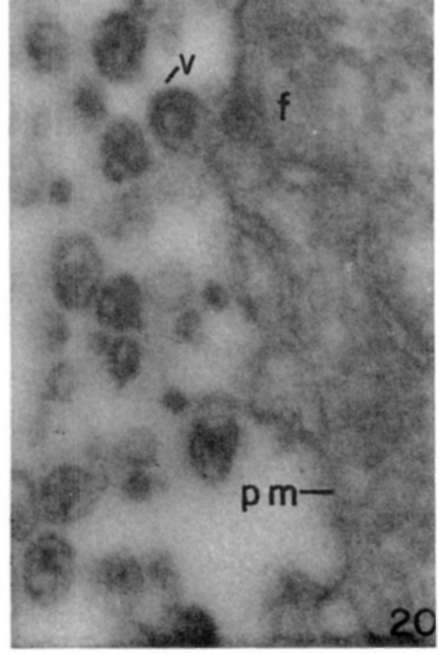
Figure 20. Fine filaments (f) in peripheral cytoplasm are near where a maturing particle (v,) is broadly attached to plasma membrane (pm). X 120,000.
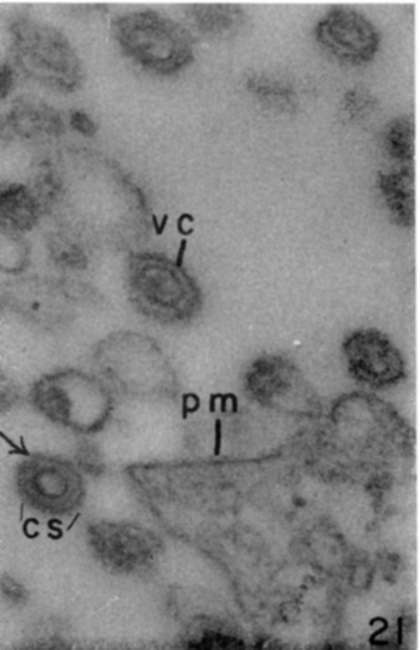
Figure 21. Viruses already detached from plasma membrane (pm) and possessing distinct coats (vc) with cross striatums (cs). Arrow indicates probable presence of pore. X 140,000.

Continue to:
My Books

