Serial Virus Passage. Part 2
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Serial Virus Passage. Part 2
Physical Properties. Electron Microscope Studies
The ultrastructure of the virus particle and its association with the leukemic tissues are being studied by Dr. A. J. Dalton of the National Cancer Institute. Observations have been made on thin sections of the virus-induced leukemic tissues and on pelletized virus concentrates prepared from these tissues. The techniques used for fixation and staining were similar to those published previously (60, 61).
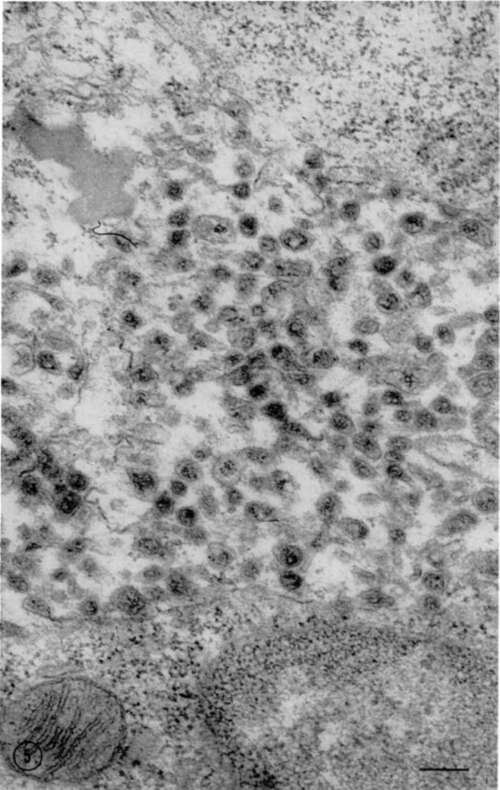
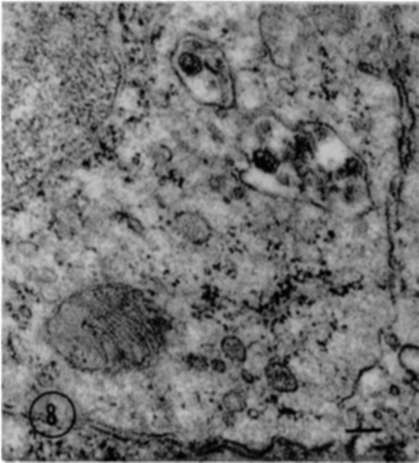
Particles were found in greatest number in an extracellular position in lymph nodes and the thymus glands. The majority of these particles were not well defined; both the nucleoid and outer membranes appeared shrunken and distorted (fig. 5). Occasionally particles were found in the cytoplasm, usually within vacuoles which possessed discrete membranes (figs. 6 and 8). Only rarely were individual particles encountered which appeared to be lying free in the cytoplasm, and these consistently exhibited good internal structure (fig. 7).
Characteristically the extracellular particles possessed a central nucleoid with an average diameter of 65 mµ. The periphery of this nucleoid was more electron dense than its central portion. Two concentric outer membranes were present, the inner with a diameter of approximately 85 mµ, and the outer with a diameter of approximately 100 mµ (fig. 9).
More comprehensive electron microscope studies are now in progress to elucidate further the ultrastructure of the active agent.
Thermal Stability
Sarcoma 37 and the virus-induced leukemic tissues maintain their capacity to produce tumors on implantation and their efficiency as virus source materials when sealed and stored at the temperature of dry ice.
Lyophilization of the particulate materials does not significantly reduce the biological activity of the virus. Data obtained from one experiment indicate no apparent difference in the activity of lyophilized material relative to a frozen aliquot of the same virus preparation.
The leukemia virus prepared in 0.05 m pH. 6.8 sodium citrate buffer is stable for at least 90 minutes at 37° C. However, the leukemogenic activity of the virus in the same buffer or in 0.12 m NaCl is completely destroyed within 30 minutes when heated at 56° C.
Summary And Conclusions
Presented herein are a summary of studies with the known viruses that induce leukemia in murine animals and the results of studies with a virus originally recovered from a connective-tissue neoplasm, Sarcoma 37, but which, when extracted from this tumor, evoked leukemia in other susceptible hosts.
Table 6. Properties Of The Murine Leukemia Viruses
Host sensitivity | ||||||
Age | Strain | |||||
Investigator | Origin | Particle size (mµ) | Suckling (percent) | Adult (percent) | Neoplastic cell (type) | |
Gross | AK, C58 (spontaneous leukemia) | 60-150 (3 types) | 62-100 | 44-60 | C3H, C3Hf (Bittner) | Lymphoid |
Schoolman and Schwartz | Swiss and C3HeB (spontaneous leukemia) | - | - | 15-100 | Swiss and C3H | Lymphoid (sarcoma) |
Graffi | Sarcoma I, Sarcoma II, SOV 16, Ehrlich carcinoma, Sarcoma 37 | 70 | 70 | 25 | Agnes Bluhm, sg, db, (rats) | Myeloid |
Friend | Ehrlich carcinoma | 78 | - | 85-100 | Swiss, DBA/2 | Reticulum cell with erythroblastosis |
Kaplan | C57BL/Ka (X-irradiated) | - | 69 | - | C57BL/Ka, compatible F1 hybrids | Lymphoid |
Moloney | Sarcoma 37 | 90-110 (extracellular) | 100 | 100 | BALB/c, C3H, C3Hf, I, C57BL, DBA/2. RIII, Swiss, A/LN, Fi hybrids, (rats) | Lymphoid |
Table 6 briefly summarizes certain of the biological and physical properties of the virus recovered from Sarcoma 37 and relates these to analogous observations with the other murine leukemia agents.
The present leukemia virus differs from two other agents recovered from solid tumors primarily by the pathology of the resultant neoplasm. It induces a generalized leukemia characterized by a lymphoid type of neoplastic cell. The agent of Graffi induces tumors in mice and rats that are characterized by a myeloid type of neoplastic cell. The Friend virus induces a leukemia in adult Swiss and DBA/2 mice which has been interpreted as a reticulum-cell disease with nonmalignant erythroblastosis.
Among the group of leukemia viruses recovered from spontaneous lymphocytic neoplasms, the Schoolman and Schwartz agent is, as stated earlier, distinguished by (1) the very short latent period for its action, (2) the formation of a lymphosarcoma-like mass involving the mesentery and (3) its recovery from the brains but not from tumors of infected mice.
The Gross agent, the first to be recovered from a naturally occurring lymphocytic leukemia, is apparently highly strain specific; it is most effective in C3H mice of the Bittner substrain. Although the lymphoid neoplasm produced by the virus is similar to the spontaneous disease, fibroinyxosarcomas and carcinomas of the parotid gland develop in mice inoculated at birth with the same cell-free preparations. This indicates the presence of more than one agent in the leukemic tissue filtrates. Of significance is the recent development, by Gross, of a highly active virus passage material effective in weanling and young adult C3H mice. Mice older than 1 day at the time of virus inoculation develop leukemia but not parotid-gland tumors.
The direct relationship of the virus described herein to the virus of Gross and other murine leukemia agents must await the outcome of immunological, physical, and biochemical studies. However, as indicated in table 6, the properties which characterize the virus are: (1) Infection is not limited to the very young animal or mice of the same genetic line; (2) lack of strict species specificity is indicated by the viral induction of lymphocytic leukemia in rats; (3) no tumors other than those of the generalized lymphocytic type have been observed, regardless of the strain or age of the test animals; and (4) virus concentrates induce leukemia in 100 percent of the inoculated animals within a relatively short period.
Thus, there is available for laboratory investigations a murine leukemia virus of high biological activity which is not age specific or highly strain or species specific.
Plate 1
Figure 1. Leukemic mice of various strains which had received virus shortly after birth. Above, normal uninoculated BALB/c mouse. Lower left, mouse of strain I. Center, mouse of strain C3Hf. Lower right, mouse of strain BALB/c. Note enlargement of spleen, liver, and thymus gland in all leukemic mice. In addition, note enlargement of mesenteric lymph node in inoculated BALB/c mouse.

Plate 2
Figure 2. Sprague-Dawley nits. Left, normal uninoculated rat. Right, rat inoculated with mouse-derived virus shortly after birth and killed at 9 weeks of age. Note enlarged spleen and thymus gland in leukemic rat.
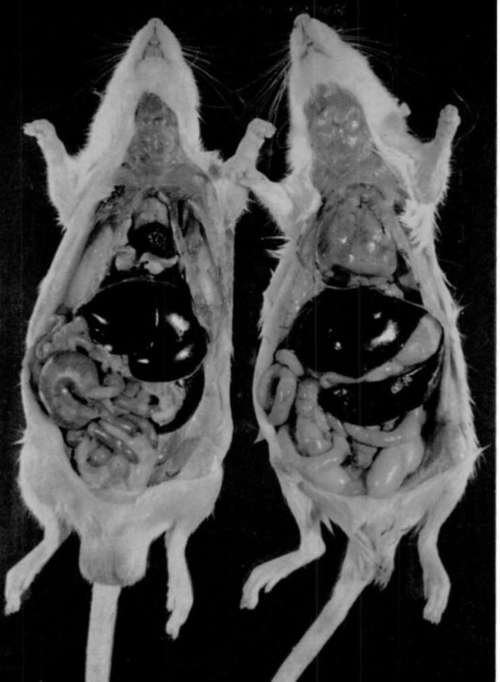
Plate 3
Figure 3. Sprague-Dawley rat inoculated with mouse-derived virus shortly after birth and killed at 10 weeks of age. Neoplastic cells in tracheobronchial lymph node. Note basophilic nuclei and narrow rim of cytoplasm. Hematoxylin and eosin. x 780.
Figure 4. Same rat. Pancreatic lymph node, normal architecture destroyed by infiltration with neoplastic cells that have extended from lymph node to infiltrate adjacent pancreatic tissue. Hematoxylin and eosin. x 150.
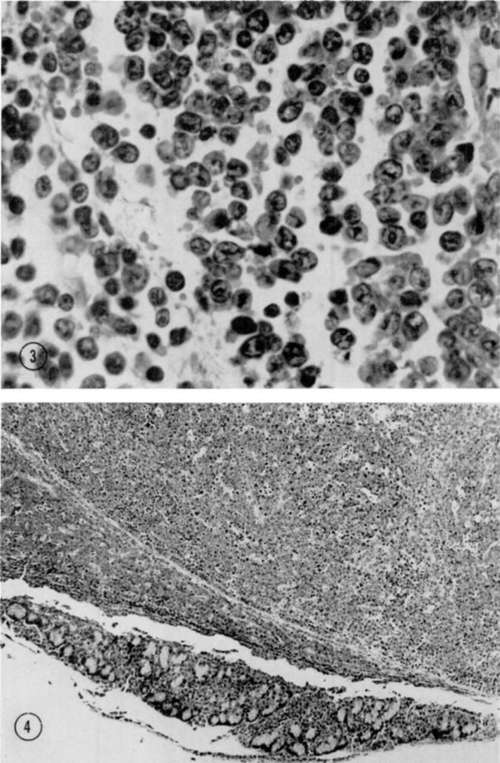
Plate 4
Figure 5. Electron micrograph of part of section through lymph node of leukemic mouse. Numerous particles varying in size and state of preservation are present in extracellular position. Horizontal line represents 0.1µ. Chrome osmium fixation. Modified lead hydroxide stain. X 70,000.
Horizontal line in each figure represents 0.1µ.
Plate 5
Figure 6. Electron micrograph of part of cell in mouse leukemic lymph node, showing intracellular particles. Majority of particles are within vacuoles delineated by distinct membrane. A few extracellular particles are present at right of figure. Chrome osmium fixation. Modified lead hydroxide stain. X 70,000.
Figure 7. From thymus of leukemic mouse. Shows solitary particle (arrow) apparently lying free in cytoplasm. This particle appears well preserved. Chrome osmium fixation. Modified lead hydroxide stain. X 70,000.
Figure 8. Shows portion of cell from lymph node of leukemic mouse. Cytoplasmic vacuoles are present containing several particles. Chrome osmium fixation. Monobasic lead acetate stain. X 52,000.
Figure 9. Shows single extracellular particle from lymph node of leukemic mouse. Details of internal structure are evident, including electron dense periphery of central nucleoid and relationship of two concentric peripheral membranes. Chrome osmium fixation. Modified lead hydroxide stain. X 157,000.



Continue to:
My Books

