2. Proteins and Amino-acids
Description
This section is from the book "Chemistry Of Enzymes In Cancer", by Franz Bergel. Also available from Amazon: Chemistry Of Enzymes In Cancer.
2. Proteins and Amino-acids
This is not the place to discuss details of protein biosynthesis and degradation, unless it has some bearing on the theme of this monograph. On the one side, proteins as histones and protamines are intimately connected with DNA in form of nucleoproteins, on the other RNA and nucleotides are considered to be indirectly or directly responsible for the coding and production of polypeptides, including those of which apoenzymes are made (EFS). Adding to this the derivation of amino-acids, such as alanine, aspartic and glutamic acid, etc., by transamination from corresponding keto-acids of the Krebs cycle and the requirement of ATP, produced by glycolysis and respiratory metabolism, as a source of energy for peptide bond formation, the close inter-relation between the cell chemistry of proteins, nucleic acids, carbohydrates and their derivatives and of lipids is quite obvious.22,83 266 On the whole, protein synthesis in tumors, just as in rapidly growing embryonic tissue is high, as demonstrated among other experimental approaches, by amino-acid incorporation. This might happen even at the expense of the tumor-bearing host. Busch and Greene 82 claimed the surprising preferential incorporation of a whole serum protein fraction into the Walker tumor as compared with other organs of the Walker rat. In the case of single amino-acids, Weisberger and Levine 262 noted, when using labelled compounds, that human leucocytes of acute and chronic myelogenous leukemia had a greater avidity for glycine and cystine (or cysteine) than normal leucocytes, although Baldini and Sacchetti21 observed that cysteine and cystine could protect healthy human bone marrow in tissue culture against cellular degeneration. De Lamirande and his group165 found a significant increase of cystine-cysteine in primary tumors of the rat liver which in difference to that noted in regenerating liver appeared to be irreversible. This together with Weisberger's 262 results with leucocytes is interesting in view of the relation between sulfhydryl groups and cell division (Rapkine,208 Chalkley,67 Mazia m-178 and Jensen141). There may be also some connection between these findings and the deficiency in cysteine degrading enzymes (or their coenzymes) in tumors as mentioned before and to be discussed later. Roberts et al. 217 studying the pool of free amino-acids and observing a specific pattern in the quantity of such amino-acids in a number of tumors (reminiscent of Greenstein's postulate of an enzyme tumor pattern) , found that proliferating neoplastic cells were taking up glutamine, at a high rate although its level in cells and fluids of tumor tissues was low.
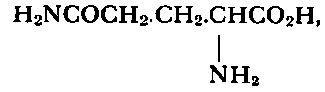
The question arises again whether all this is due to an imbalance between anabolising and catabolising enzymes, governing protein, peptide and amino-acid formation and degradation. So far most of the results have been obtained with Novikoff hepatoma, representing perhaps rather an extreme case and quoted before in connection with degrading enzymes of the nucleic acid series. While the Montreal group reported complete deficiency in glutamic acid dehydrogenase8 (see Fig. 13), which is seemingly increased in leukemic leucocytes,246 Auerbach and Waisman 16 found that a whole series of other amino-acid degrading enzymes were absent in this hepatoma: tryptophane peroxidase-oxidase, phenylalanine hydroxylase, p-hydroxyphenylpyruvic acid oxidase, histi-dase, tyrosine transaminase, threonine and serine dehydrases and cysteine desulfhydrase 229 (Fig. 13).
The activity levels of phenylalanine transaminase, arginase, glutamine synthetase (Fig. 13) and cathepsin 6 were diminished in this liver tumor.
Thus on the whole, a similar situation prevails as in the field of nucleolytic and purine and pyrimidine degrading enzymes. Although the glutamine synthetase has to be considered as an anabolic biocatalyst (providing this amino-acid amide perhaps as co-factor in de novo purine synthesis68) and cathepsin could function as such, the overall situation is again that of an apparent deletion of catabolic enzymes, at least in the Novikoff hepatoma. This loss, bringing about a preservation of amino-acids, could explain the tendency toward increased protein synthesis, whereby certain apoenzymes seem to escape selectively the general anabolic activity. Whether the tumors, to ask the old question again, are deficient in EFS, holo- or co-enzymes or whether some enzymes are temporarily or permanently inactivated, possibly due to changes in sub-cellular particles such as microsomes, mitochondria, etc., can be answered at least in specified cases. It has been mentioned before that lack of 'cystine desulfurase' activity is caused by absence of the coenzymes, pyridoxal phosphate and DPNH,231 at least in the mouse tumor investigated. The situation may be somewhat different in the Novikoff hepatoma where Allard et al.6 found that of two transaminases, both dependent on pyridoxal phosphate, one was detectable whereas the other was absent. Moreover, Auerbach and Waisman 16 again with Novikoff hepatoma were unable to induce an increase in the levels of tryptophane peroxidase, tyrosine transaminase and threonine dehydrase by application of the respective substrates which in the case of tryptophane peroxidase and less so of the transaminase, produced such an adaptive response in the adjacent healthy liver tissue. This means that the deficiencies noted go beyond the coenzyme and even holoenzyme levels and may involve the corresponding EFS.
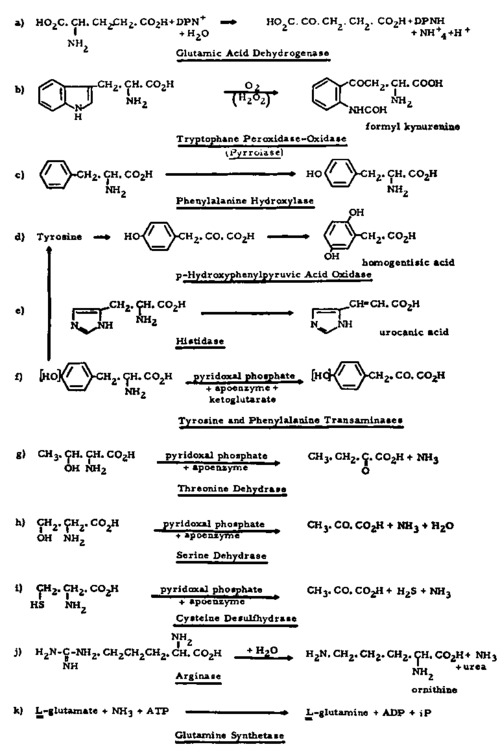
Fig. 13. The degradation of amino-acids catalyzed by catabolic enzymes.
Continue to:
My Books

