Enzymes And Co-Factors Of Interest
Description
This section is from the book "Chemistry Of Enzymes In Cancer", by Franz Bergel. Also available from Amazon: Chemistry Of Enzymes In Cancer.
Enzymes And Co-Factors Of Interest
It proposed to discuss this matter under the sub headings of metabolism of nucleic acids (standing for polynucleotides, nucleotides, nucleosides, purines and pyrimidines), of proteins and ammo acids, of carbohydrates (glycolysis, pentose shunt, Krebs cycle and oxidative phosphorylation), and of lipids; of function and synthesis of coenzymes and of the role of trace elements. It has to be borne in mind that in spite of this sub division the various metabolic pathways of all cellular constituents and their enzyme systems, as mentioned just now, are well integrated and interdependent and that they consist of catabolic (degradative) and anabolic (building up) streams (see Fig. 5). One has also constantly to think of the distribution of the enzymes in different 'compartments' of the cell, nucleus, nucleoli, mitochondria, lysosomes, microsomes, endometrium, cell membrane, etc. and of the sometimes profound differences between the effect of enzymes attached to sub cellular structures and those liberated from injured cells and freely dissolved in buffered solutions. It is understood that in the framework of this monograph only a selection of examples will be given.
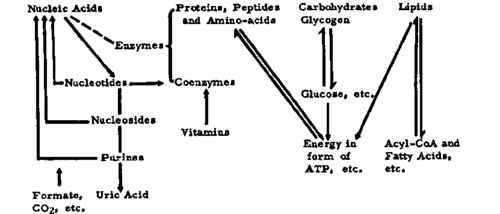
Fig 5. Catabolic ↓ and anabolic ↑pathways.
1. Nucleic Acids
"Everything comes back finally to nucleic acid." This is how Todd, during the first Jephcott Lecture,242 summarised his ideas on the present state of the two hypotheses attempting an explanation of the origins of cancer, the postulates of somatic mutation and of virus effects. It is therefore not surprising that nucleic acids and their building units are placed by so many workers in the center of their investigations and speculations. So that some picture of the complicated events, which in some cases so far have only been observed with micro organisms, can be obtained, a table on the nucleic acid, etc. metabolism is presented, laying emphasis on the anabolic and catabolic enzyme systems involved (Fig. 6). No claim is made to its completeness. On the left the principal synthesising or anabolic enzymes and on the right the degrading or catabolic enzymes are arranged against larger molecules at the top and smaller molecules at the bottom. One could draw in one's mind arrows which would point upwards for anabolism and downwards for catabolism, but this would be an oversimplification of the actual events under in vivo conditions. At the same time it is undesirable to lose oneself in the complex details. Nevertheless, another diagram 29 in Figure 7 shows the interconversion of purine derivatives, forming a kind of 'cycle.' It has been added to help in the understanding of some of the points raised below. The fact which should not be forgotten is that, according to present day knowledge, the only purine derivative formed de novo by biosynthesis is inosinic acid (see also Fig. 6). It appears to be the mother substance of all other purines and their derivatives which, of course, may enter the cycle also in form of preformed material (from food and polynucleotide breakdown products).
As mentioned above, the role of DNA and RNA in the life of cells is a very crucial one. How do these macromolecules, in combination with proteins (histones and protamines), the smaller building units and the related enzymes behave in tumor cells in comparison with those in embryonic, normal regenerating, proliferating (intestinal and bone marrow), resting and benign tumor cells? Are there any differences detectable between all these compounds in various cell types, differences reflecting particularly on the 'unrestricted growth' of the malignant tissue, and of course linked up with events concerning other groups of cellular constituents as depicted in Figure 5? DNA which carries most of the markers of the genotypical and phenotypical properties of the cell and acts the part, so to speak, of an information center is metabolically" very stable (unless changed by mutagenesis including, maybe, carcinogenesis) and only increases in amount by anabolic processes accompanying some phases of cell division or mitosis. This is illustrated by a diagram (Fig. 8) drawn up by Walker and Yates.247 Another scheme was put forward by Lajtha181 which again depicts the same phenomenon (Fig. 9). RNA on the other hand, is less 'metabolically stable' in rapidly growing cells and possibly more subject to renewal in resting cells.

Fig. 6. Nucleic acid, etc. metabolism (enzymes in italics). Sugars: RNA-ribose from pentose shunt, DNA-deoxyribose from?.

Fig. 7. "Purine cycle". (From Bergel et al: Ciba Foundation Symposium." Courtesy of J. and A. Churchill, Ltd., London).

Fig. 8. Nuclear components of dividing cells: O DNA, o polynucleotides and mononucleotides, P=prophase, M=metaphase, T=telephase of cell division. (From Walker and Yates: Proc. Roy. Soc. BS*' Courtesy of The Royal Society, London.).
Continue to:
My Books

