Examples Of Enzymes And Coenzymes In Carcino-Chemotherapy
Description
This section is from the book "Chemistry Of Enzymes In Cancer", by Franz Bergel. Also available from Amazon: Chemistry Of Enzymes In Cancer.
Examples Of Enzymes And Coenzymes In Carcino-Chemotherapy
In the following, a number of selected investigations are reported, together with the rationale why they were undertaken. They were mostly carried out with animals, and with the exception of one case, no clinical trials have been undertaken yet. This means that the final proof of their significance in human cancer has still to come.
1. Xanthine Oxidase
It has been mentioned before, either that certain tumors had low activity levels of xanthine oxidase, or that such activity was completely absent as in the Novikoff hepatoma.156 This state of affairs, it will be remembered, was also discussed by Bennett et al.2B But the matter goes further back than the report by Lewin et al.178 (see also reference 29). Indeed the findings of these authors with mammary tissues of mice came a considerable while after Haddow et al.110 made the observation that xanthine oxidase from cow's milk inhibited the growth of spontaneous mammary carcinoma in C3H and C+ mice, and even longer after Haddow115 had reported that xanthopterin, a naturally occurring pteridine, induced in rats and other rodents a transient kidney hypertrophy, probably due to stimulation of mitotic activity in the tubular epithelium. Apart from xanthopterin, other pteridines, pteridine analogues, purine analogues, etc. (see Bergel28) were tested and some of them found to produce the same phenomenon. As xanthopterin acts as a substrate and in higher concentration as an inhibitor of xanthine oxidase, Albert 4 and Timmis 238 (see Haddow et al.120) independently put forward the following hypothesis: Through inhibition of the enzyme either an accumulation of xanthopterin which is a growth-promoting substance could cause a stimulation of cell division, or if the concentration of xanthine increased in the cell by not being degraded to uric acid, etc., this could have repercussions on the purine and nucleotide pool and finally on nucleic acid synthesis. Following this idea, the in vitro inhibition of xanthine oxidase from cow's milk by the substances tested for their production of kidney hypertrophy was investigated,36 with the result that in some instances there was a correlation between the chemical and biological effects, in others there was none. However, in view of the speculations concerning the enzyme, and, as it was stated before, in view of the carcinogenic effects of long-term administration of xanthine,29 a purification of xanthine oxidase from cow's milk was undertaken (see Avis et al.11) which led to a crystalline preparation. Continuation of this research,18 together with contributions from many other places,80 have given some insight into the chemical constitution of the enzyme. It has a molecular weight of about 300,000 and contains in addition to two molecules of FAD, 8 atoms of iron and 1-2 atoms of molybdenum. These co-factors are very strongly held by the apoenzyme, and their role in the catalytic mechanism is slowly unravelled (see Fridovich and Handler100 and Bray et al.64). Xanthine oxidase does not only catalyse the oxidation of hypoxanthine to xanthine and xanthine to uric acid, but also that of other purines and pteridines and of a great number of aldehydes. All this has to be taken into account when considering the following studies121: Female C3H and C+ mice, carrying the Bittner milk factor,38 develop after mating spontaneous mammary carcinomas. The animals were treated by i.p. injections of highly purified and active bovine xanthine oxidase (XO) (5 mg./mouse/daily) against a control group receiving saline injections. No cure was achieved but, as can be seen from Figure 19, an arrest of the growth rate of the tumors which were measured two-dimensionally with calipers was produced. The impression was gained that on cessation of the XO injections, the tumors regained the original growth rate prior to the first treatment. As the enzyme preparation came from a different species and contained rather large molecules, the questions arose whether the observed anti-tumor effect was of a non-specific nature and whether the 'giant' enzyme was taken up at all by the mouse tissues if not by the cells. This was in part answered by the injection into non-tumor bearing mice of 5 mg. active or inactive XO (inactivated by U. V. radiation) or the supernatant of 5 mg. of heatdenatured enzyme or 10 mg. of crystalline bovine serum albumin. When the livers of these animals were analyzed aerobically for XO-activity, it was found (see Fig. 20) that only the active preparation caused its marked and temporary increase, the inactive sample appeared to cause a less pronounced one after a lag period (possibly for reactivation), while the albumin, supernatant or both had no effect. Moreover, when increasing amounts of XO were injected (see Fig. 21), the rise in the XO-activity levels was practically proportional to the dose up to about 0.1 units of XO (for the definition of units, see Avis et al.17). However, even more encouraging results were obtained when the XO-levels of tumors and livers of treated and untreated (control) animals were compared (Table I). Apart from the livers, the tumor tissue of the animals injected with the enzyme preparation showed a significantly higher XO-activity than of those which were given only saline injections. There remain still the problems of cell penetration, the underlying mechanism and of immunological properties. So far no evidence is available that the macro-molecules have entered the cell, and it is not unlikely that they accumulate in the intercellular fluid. The mode of action could amount to a restitution therapy, the injected XO making good the deficiency in catabolism, as discussed before. Further work on the immunological characteristics of XO from cow's milk is imperative. It can be stated that in preliminary experiments31 an anti-serum, produced in rabbits, acted as a moderately effective inhibitor of the bovine enzyme in vitro (see Cinader73). It is not impossible that the mouse with its weaker immuno-response reacts less violently to the injections of bovine preparations than other species. The prospects of utilizing these results for human medicine are somewhat clouded by preliminary findings that XO-activity in normal and cancerous breast tissue from women seems to be equally very low. Whether the human tissue in its nucleic acid catabolism bypasses the hypoxanthine and xanthine stages and disposes of these purines by oxidising inosinic acid or inosine (hypoxanthine ribo-tide and riboside, see Fig. 7) to uric acid derivatives will have to form the subject of further investigations, together with that of a great number of other problems.

Fig. 19. Growth curves of spontaneous mammary tumors of mice treated with xanthine oxidase against control groups ( 18 C3H and C+ mice on saline injections; oo 18 animals treated with 5 mg. xanthine oxidase daily. Relative tumor surface surface of averaged tumors at each day x 100/surface of averaged tumors at time zero.) (From Haddow et al.: Nature."' Courtesy of Macmillan and Co. Ltd., London.).

Fig. 20. Effect of single injections on the levels of liver xanthine oxidase in non-tumor bearing mice, (oo active XO, 5 mg.; inactive XO; aa bovine serum albumin, 10 mg. or supernatant from heat denatured enzyme or both. The first point is the mean of determinations of 12 mouse livers, the others of 3 from treated animals.) (From Haddow et al.: Nature.1*1 Courtesy of Macmillan and Co. Ltd., London.).
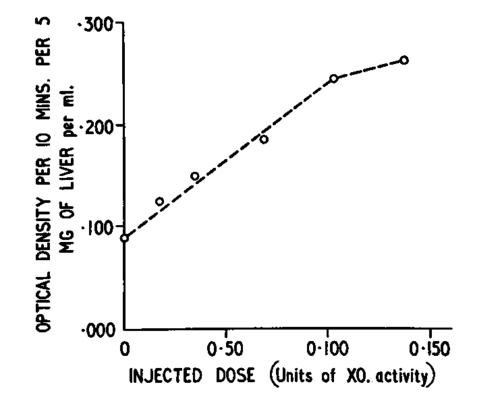
Fig. 21. Relation between liver xanthine oxidase activity in mouse livers of non-tumor bearing animals (the first point is the mean of determinations of 12 mouse livers, the following, all 4 hr. after injection, of 2) and dose level of injected XO. (Form Haddow et al.: Nature. Courtesy of Macmillan and Co. Ltd., London.).
Table 1
Effects of xanthine oxidase injections on tumor-bearing animals (some-tumor-bearing mice, from Figure 19, were killed after fifteen daily injections of XO or saline. The tumor tissue and the livers were used for XO-assays; activity is expressed in terms of uric acid formed per mgm. of fresh tissue). (From Haddow et al.: Nature.
Courtesy of Macmillan and Co. Ltd., London.)
No. of Animals | 10^6µ moles/min./mgm. Mean Xanthine Oxidase Activity | ||
Tumour | Liver | ||
Treated Controls | 4 6 | 24.4 10.0 | 40.0 14.3 |
Continue to:
My Books

