The Function Of Cytidine Coenzymes. Part 5
Description
This section is from the book "The Scientific Contributions Of The Ben May Laboratory For Cancer Research", by The University of Chicago. Also available from Amazon: The Scientific Contributions Of The Ben May Laboratory For Cancer Research.
The Function Of Cytidine Coenzymes. Part 5
The incubation mixture used in the experiments on the enzymatic synthesis of CDP-choline and CDP-ethanolamine contained the following components in a volume of 2.0 ml.: 20 pmoles of MgCl2, 100 pmoles of phosphate buffer, pH 7.4, 10 pmoles of cysteine, 20 µmoles of ATP, 0.4 µmole of CTP, and 20 pmoles of P-choline-P32 or P-ethanolamine-P32. 1.0 ml. of enzyme was added, and the tubes were incubated for 1 hour at 37°.
The reaction was stopped by the addition of 5 ml. of 66 per cent ethanol at room temperature. The precipitate of denatured protein was removed by centrifugation and washed with 10 ml. of water. The combined extracts were diluted to a volume of 60 ml. and chromatographed on a column of Dowex 1 formate, the formic acid system being used. The fractions which were collected were then analyzed for radioactivity and for absorb-ancy at 280 mp. The results are shown in Fig. 6. In the experiment with P-ethanolamine-P32 (lower portion, Fig. 6), the unchanged P-ethanolamine was found in Tubes 11 to 20. About 50 per cent of the added CTP was recovered as CDP-ethanolamine in Tubes 30 to 36. In the experiment with P-choline-P32(upper portion, Fig. 6), about 70 per cent of the CTP was recovered as CDP-choline in Tubes 22 to 29.
The enzymatically synthesized CDP-choline was identical with the synthetic product, as judged by rechromatography on ion exchange resin (ammonium formate system), paper chromatography, absorption spectrum, and biological activity. The enzymatically synthesized CDP-choline was converted to lecithin by rat liver particles in yield identical with that observed with the synthetic product.
Reversibility of PC-qjtidyl Transferase-The enzymatic synthesis of CDP-choline shown in Reaction a should be readily reversible, since it is closely similar to the formation of DPN from ATP and nicotinamide ribo-tide which has been found by Kornberg (17) to have an equilibrium constant of about 0.4. PC-cytidyl transferase should therefore catalyze not only the synthesis of CDP-choline from CTP and P-choline, but also pyro-phosphorolytic cleavage of CDP-choline to yield CTP and P-choline. In the experiment presented in Table III, the addition of small amounts of pyrophosphate to the enzyme system markedly reduced the over-all incorporation of P-choline into lecithin. This effect was interpreted as indicating that the equilibrium of Reaction a had been displaced to the left.
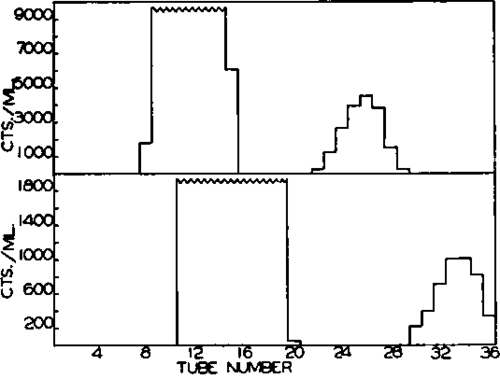
Fig. 8. Chromatographic separation of enzymatically synthesized CDP-choline (upper portion) and CDP-ethanolamine (lower portion). Details of the experiment are given in the text.
The pyrophosphorolysis of CDP-choline labeled with choline-1,2-CM was studied in the experiment described in Table VII. After incubation for 15 minutes at 37° under the conditions described, the enzymatic reaction was stopped by immersing the tubes in a boiling water bath for 6 minutes. The suspension was centrifuged and 0.50 ml. of the supernatant fluid assayed for P-choline-1,2-C14. Carrier P-choline (20 pmoles) was added and the pH adjusted to 4.7 with acetate buffer. The P-choline was then quantitatively hydrolyzed to choline by the addition of acid phosphatase from semen, and the choline liberated was recovered as the crystalline reineckate and counted. From the total radioactivity of the choline reineckate and the specific activity of the original CDP-choline, the amount of P-choline formed by the PC-cytidyl transferase reaction could be calculated. The results are given in Table VII. A significant amount of P-choline is formed by the action of purely hydrolytic enzymes, but this value is increased almost 6-fold by the addition of inorganic pyrophosphate, indicating the reversibility of Reaction a. Inorganic orthophosphate is without effect on the reaction.
Table VII. Pyrophotphorolytot Of Cdp-Choline
Tube No. | CDP-choline split | |
mµmoles | ||
1 | No pyrophosphate added | 23.8 |
2 | 5 pmoles pyrophosphate added | 123 |
The enzyme system for the pyrophosphorolytic cleavage of CDP-choline contained the following reagents: 20 pmoles of MgCl2, SO pmoles of Tris buffer of pH 7.4, 0.6 pmole of CDP-choline labeled with choline-1,2-C14 (44,000 c.p.m. per micromole), 6 pmoles of cysteine, and 0.6 ml. of the 0 to 40 ammonium sulfate fraction similar to that used in the experiment in Fig. 6. The tubes were incubated at 37 degrees for 16 minutes.
Distribution of PC-cytidyl and PE-cytidyl Transferases-These enzymes are widely distributed in nature, being found in the liver, brain, heart, and kidney of the rat, in the fiver of the guinea pig and hog, in several strains of yeast, and also in carrot root. A detailed study of the properties of the PC-cytidyl transferase of guinea pig liver has been made and will be published in another paper.
PC-glyceride and PE-glyceride Transferases-Reaction b, Fig. 3, may be regarded as essentially the transfer of the P-choline portion of CDP-choline to the free hydroxyl group of a D-a,β-diglyceride. The enzyme catalyzing this reaction will be described as PC-glyceride transferase; the analogous but separate enzyme catalyzing the formation of phosphatidylethanolamine from CDP-ethanolamine will be described as PE-glyceride transferase. Evidence that two distinct enzymes are required for these reactions has been given in Table V.
Properties of PC-glyceride Transferase-Added magnesium or manganese ions are necessary for PC-glyceride transferase activity (Fig. 7), just as for the over-all reaction sequence by which labeled P-choline is incorporated into lecithin (Fig. 2). Calcium ions inhibit PC-glyceride transferase even in the presence of magnesium ions, whereas PC-cytidyl transferase is unaffected by calcium. The inhibition of the over-all system by calcium ions (4) is therefore localized in the PC-glyceride step.
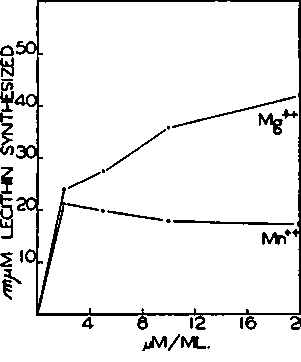
Fig. 7. Requirement of divalent cation for PC-glyceride transferase activity. Each tube contained 0.10 µmole of CDP-choline-1,2-C14 (65,000 c.p.m. per micromole), 100 µmoles of phosphate buffer of pH 7.4, and 0.50 ml. of rat liver particles in a final volume of 1.0 ml. Magnesium or manganese ions were added as shown. Incubation was for 1 hour at 37°.
Table VIII. Nucleotide Specificity Of Pc-Glyceride Transferase Of Rat Liver And Yeast
P-choline incorporated into lecithin, mpmoles | |||
Tube No. | Liver enxyme | Yeast enzyme | |
1 | UDP-choline | 0.7 | 0.3 |
2 | GDP-choline | 0.0 | 0.0 |
3 | ADP-choline | 0.0 | 0.0 |
4 | CDP-choline | 64.8 | 84.1 |
Each tube contained 40 pmoles of MgCl2, 100 pmoles of Tris buffer, pH 7.4, 20 pmoles of cysteine, 100 pmoles of KF, and 0.6 ml. of rat liver particles or crude di-alyzed autolysate of brewers' yeast as indicated. 0.20 pmole of labeled nucleotide of identical specific activity (55,000 c.p.m. per micromole) was added as shown. The tubes were incubated for 1 hour at 37°. The final volume was 2.0 ml.
PC-glyceride transferase is highly specific for CDP-choline. When tested with synthetic UDP-choline, GDP-choline, ADP-choline, and CDPcholine (Table VIII), only CDP-choline was found to be active. A similar narrow specificity was found with both the liver and the yeast enzymes.
Requirement for D-a,β-Diglyceride-Preparations of particles from the livers of animals of several species convert CDP-choline to lecithin in the absence of added diglyceride. These enzyme preparations are rich in lipide which is bound tenaciously, and the addition of suspensions of lipides of various types suspected to be precursors of the a,β-diglyceride moiety of lecithin was without consistent effect. Similar results were also obtained with enzymes from yeast.
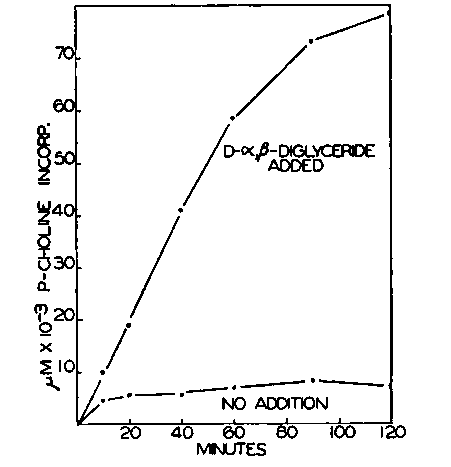
Fig. 8. Requirement of d-a.β-diglyceride for conversion of CDP-choline to lecithin. Each tube contained 20 µmoles of MgCl2, 50 pmoles of Tris buffer of pH 7.4, 0.2 pmole of CDP-choline (58,000 c.p.m. per micromole), 10 pmoles of cysteine, 5 mg. of Tween 20, and 0.2 ml. of rat liver particles in a final volume of 1.0 ml. 3 mg. of D-a,β-diglyceride prepared from purified egg lecithin were added as indicated. The diglyceride was added as an emulsion in the Tween 20; the control received Tween 20 only. The incubation at 37° was varied as shown.
Continue to:
My Books

