Studies On Broken Cell Preparations. Part 2
Description
This section is from the book "Symposium Phenomena Of The Tumor Viruses", by U.S. Dept. of Health. Also available from Amazon: Tumor Suppressing Viruses, Genes, and Drugs: Innovative Cancer Therapy Approaches.
Studies On Broken Cell Preparations. Part 2
Despite the obscurity of the initial organelle involvement, the subsequent course of the process is clear in the progression of forms from the viroplast to the vacuole, as already described. It is of much interest to find that some aspects of the progression resemble an expression of altered potentialities with which the myeloblast was initially endowed. The findings strongly suggest that the virus enters and multiplies in structures which are the precursors of the granule. In the process the organelle retains, in part, the capacity to synthesize adenosinetriphosphatase but is subsequently destroyed and, with disintegration, is converted to the vacuole, a structure basically the normal eliminative organelle of the phagocytic cell of which the myeloblast is the precursor.
Another aberration of the natural potential of the cell is the capacity of the infected myeloblast for continued proliferation. Myeloblasts of normal bone marrow succumb after brief intervals in tissue culture (24), but the virus-associated cell proliferates indefinitely at exponential growth rates under the same conditions (23). The mechanism by which the virus confers this property on the cell is not known. There has been no evidence whatever that the nucleus is directly affected. In effect, it appears to be the influence of the agent to block all maturative processes, and for practical purposes, the cell continues to express its primitive capacity to multiply. On one point there is no doubt. It is clear that the processes of virus multiplication do not interfere with those of cell reproduction, which are carried through in an adequate growth medium with the derivation of daughter cells like the parent, each with its complement of organelles including the virus-associated structures. Seemingly, the cell is able to maintain the agent essentially outside the cytoplasm in well-walled structures. There has been no evidence of intracellular rupture of virus-containing organelles, and virus particles are not seen free in the cytoplasm. Except in rare instances, in figure 6, the virus, although multiplying rapidly as indicated by the rate of release in culture (28), does not accumulate in the myeloblast. This is in great contrast to the findings with reticular cells and macrophages in the diseased chicken which may contain large amounts of the agent.
In its various expressions of neoplastic response to the etiologic virus, the myeloblast of avian leukemia is thus far unique. This is evident in the well-defined specific ultrastructures which clearly indicate that the processes of cell response are organized and incorporated, for the most part, in structures and mechanisms normal to the myeloblast. As already mentioned, this does not occur in other cells invaded by the agent in the diseased chicken. That the essence of response is related to the characteristics of the myeloblast as such, rather than to a peculiar influence of the virus, is indicated by the radically different response of the erythroblast in avian erythroblastic leukemia, to be considered in another report (48). Although the virus of erythroblastosis is immunologically closely related to the agent of myeloblastosis (22), the response and behavior of the erythroblast from the diseased bird is wholly different from that of the myeloblast. A large part of this must be related to the fundamental differences in the potentials of the two host cells. It may be noted in passing that the findings with no other virus tumor cell resemble those with the myeloblast with respect to the specificity and clarity of structural involvement.
The presence of adenosinetriphosphatase in the granules of the maturing myelocytes and in those of the heterophils may well be related to the occurrence of this enzyme in the myeloblastosis virus. It seems entirely likely that the viroplasts are related in their derivation to the granules and their precursors. This suggests that the adenosinetriphosphatase may be the product of a normal cell process and that, in its development in a medium rich in the enzyme, the virus incorporates the material into its substance. It is notable that an analogous incorporation of chick-tissue antigen has already been recognized (49). Thus, it is possible that the presence of the enzyme in the virus is specific only in relation to the peculiarities of the medium in which it is synthesized. The antigenically related virus of erythroblastosis (22) contains none of the enzyme (50) nor does the cell, the erythroblast in which it is synthesized, contain adenosinetriphosphatase-rich organelles.
Plate 35
Figure 1. Myeloblast from circulating blood of diseased bird, washed in culture fluid in preparation for culture. Swollen appearance of mitochondria (M) and membrane around nucleus (N) is rather common in circulating cells and may be increased by centrifugal washing of these cells in preparation for culture in vitro. Structures marked (V) may be mitochondria which have lost their cristae by fragmentation. There are several small gray bodies (GB) and a well-defined Golgi zone (G). Relatively small size of nucleus is probably due to plane of sectioning. X 29,000.
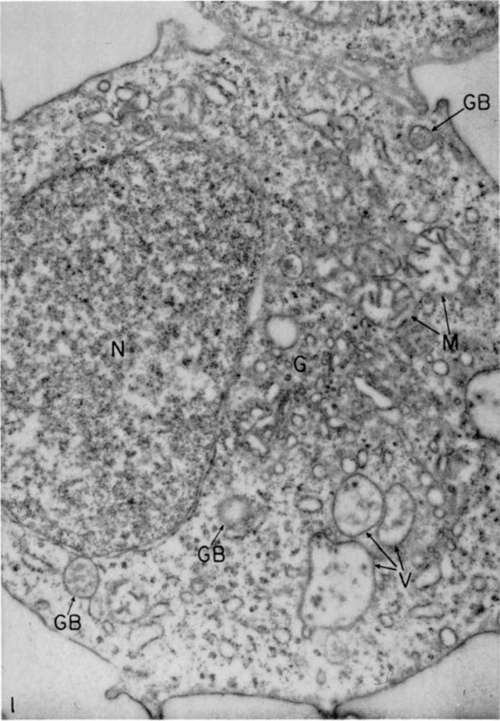
Plate 36
Figure 2. Myeloblast from same source as that of figure 1 after 2 hours in tissue culture in a growth-supporting medium. Mitochondria (M) are somewhat swollen, and two (AM) appear to be undergoing internal disorganization. Viroplasts (PL) occur frequently in sections of these cells. A virus particle (VP) lies outside cell. Structure of nucleus is shown at (N). X 48,000.
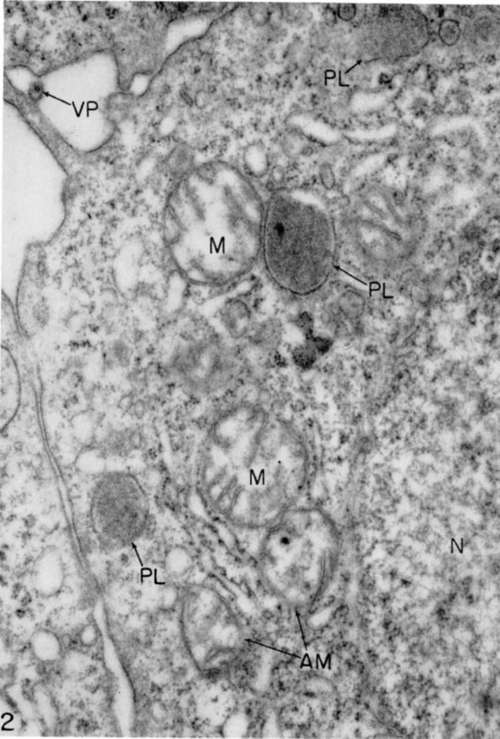
Plate 37
Figure 3. Myeloblast from same culture as that of figure 2 after 24 hours in growth-supporting medium. Section is thicker, accounting in part, at least, for greater density of the various structures. One mitochondrion of normal appearance (M) and several which seem altered (AM) are evident. While some of the appearance of alteration may be due to plane of sectioning, this does not appear to be the whole explanation. A very large viroplast (PL) contains a virus particle (VP). Nucleus is shown at (N). X 28,000.
Continue to:
My Books

