The Respiratory Metabolism. Part 6
Description
This section is from the book "The Elements Of The Science Of Nutrition", by Graham Lusk. Also available from Amazon: The Elements of the Science of Nutrition.
The Respiratory Metabolism. Part 6
These facts are brought out in Fig. 17. It should be remembered that 25 grams of glycocoll and 20 grams of alanin are each convertible into 20 grams of glucose. Leucin and tyrosin exerted only a slight effect upon the heat production. A mixture of 5.5 grams each of glycocoll, alanin, glutamic acid, and tyrosin, containing 3.46 grams of nitrogen, produced about the same specific dynamic action as 100 grams of meat which contain about 3 grams of nitrogen.
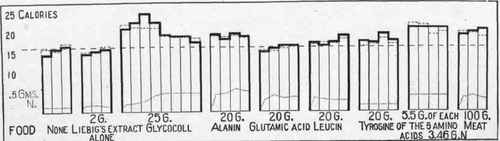
Fig. 17. - Dog II. Hourly metabolism beginning the second hour after giving amino-acids with Liebig's extract. Heavy line - calories calculated. Broken line - calories found. Dotted line - nitrogen of protein plus amino-acid eliminated.
The curve of nitrogen elimination shown in Fig. 17 does not truly represent the rapidity of the metabolism of the amino-acids. If instead of using the hourly extra nitrogen elimination after giving glycocoll or alanin one employs the "extra glucose" elimination after giving these substances to a dog with phlorhizin glycosuria, it is discovered that the maximum breakdown of the amino-acid takes place during the second hour after their ingestion. The following chart (Fig. 18) shows such experiments as accomplished by Csonka:1
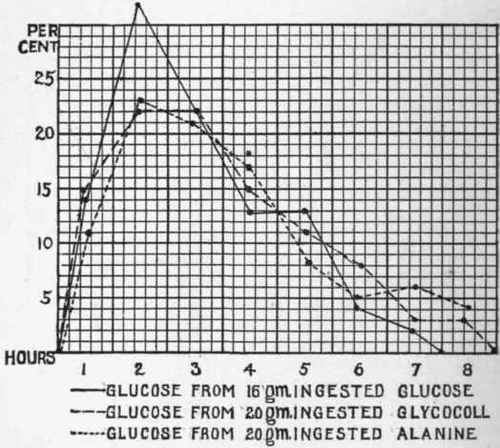
Fig. 18.
This chart shows that the rapidity of the absorption and elimination of glucose ingested in phlorhizin glycosuria is almost the same as the rapidity of the absorption, deamination, synthetic sugar production, and the elimination of such sugar after the administration of an iso-glucogenic quantity of glycocoll and a comparable quantity of alanin.
1 Csonka: "Journal of Biological Chemistry," 1915, xx, 539.
It may be added that Janney1 reports that after giving meat to a phlorhizinized dog the extra sugar appears in the urine quite as rapidly as after giving glycocoll or alanin. The rapidity of the attack of digestive enzymes upon protein must, therefore, be much greater than has hitherto been supposed.
Using the results obtained with glycocoll and alanin, Lusk2 found that the hours of the greatest heat production after the administration of these substances were coincident with the hours of their greatest metabolism. Also, it was found that the increase in metabolism after giving glycocoll and alanin together is equal to the sum of the effects produced by either alone. Furthermore, the increase of metabolism after giving 20 grams of glycocoll is twice as great as after giving 10 grams. Similar relations obtain after different quantities of alanin. This accords with Rubner's discovery that the intensity of the specific dynamic action is proportional to the quantity of protein ingested. When one compares the heat-increasing power of glycocoll and alanin upon metabolism, it is found that this power is not proportional to their respective abilities to form sugar, but rather to the number of molecules of glycollic and lactic acids which they are respectively supposed to yield on deamination.
It was found in one experiment that the entire energy content of the ingested glycocoll reappeared in the extra output of energy given off by the dog in the form of heat. The course of inquiry into this phenomenon which naturally suggests itself is whether glycocoll is without action upon the body-cells; that is, whether it merely explodes and yields heat, or whether it directly stimulates the cells, thereby raising metabolism to a higher level. This point was determined by giving glycocoll to a phlorhizinized animal. Under these circumstances there is no oxidation of the material ingested and the energy content of the glycocoll is eliminated in the urine in the form of sugar and urea. The metabolism was largely increased, notwithstanding the fact that there was no oxidation of the ingested material. Exactly the same phenomenon followed the ingestion of alanin in phlorhizin glycosuria. The ingestion of glucose was without effect even after 70 grams had been given. The cause of the specific dynamic action of glycocoll and alanin therefore lies in a chemical stimulation of the cells, causing them to metabolize more material. This confirms the older view of Voit that the action of food increases the power of the cells to metabolize.
1 Janney: "Journal of Biological Chemistry," 1915, xxii, 191.
2 Lusk: Ibid., 1915, xx, 555.
An experiment1 which shows the effect of giving 20 grams of glycocoll to a phlorhizinized dog is here reproduced:
Dog III, March 25, 1915, Experiment 104. Basal Phlorhizin Metabolism As Affected By 20 Grams Of Glycocoll In 210 C.C. Of Water At 38° Plus 1 Gram Of Liebig's Extract
Hours. | R.Q. | Calories. | ||
Indirect. | Direct. | |||
I | Basal............. | 0.733 | 23.78 | 24.53 |
2 | " | 0.716 | 23.82 | 23.84 |
Average........ | 0.724 | 23.80 | 24.18 | |
3 | Glycocoll, 20 grams | |||
4 | 0.707 | 34.21 | 32.34 | |
5 | o.745 | 31.65 | 29.47 | |
6 | 0.700 | 29.24 | 30.07 | |
7 | 0.702 | 25.99 | 26.85 | |
Average........ | 0.720 | 30.27 | 29.68 | |
Although glycocoll was not oxidized, but appeared as glucose and urea in the urine, yet there was a considerable increase in the heat production after its ingestion in phlorhizin glycosuria. It is, therefore, evident that the cause of the specific dynamic action of glycocoll is independent of the oxidation of glycocoll or the liberation of its energy content. These results, which were first presented at the International Physiological Congress held in Groningen in 1913,1 have been fully confirmed by Grafe,2 who reports that after giving 50 grams of glycocoll to a normal dog the oxygen absorption rose 77 per cent., while in man the increase was 14 per cent.
1 Lusk: Loc. cit., p. 612.
Continue to:
My Books

