Chapter II. Stems
Description
This section is from the "Histology of Medicinal Plants" book, by William Mansfield. Also see Amazon: Histology of Medicinal Plants.
Chapter II. Stems
When studying stems it should first be determined whether they were derived from monocotyledonous or dicotyledonous plants. This fact is ascertained by determining the type of the fibro-vascular bundle. See Chapter XL The next fact to determine is whether the stem is from an herb or from a woody plant. This fact is readily determined because herbaceous stems have a true epidermis, masses of collenchyma at the angles of the stem. The cortical cells contain chlorophyll, and the pith is very large. Woody stems have a corky layer, a phellogen layer, and the pith is very small except in the very young woody stems.
Having determined these facts, a study should be made of the arrangement, form, structure, color, and the cell contents of the different cells in order to determine the species of plant from which the stem was obtained.
Herbaceous Stems
The great variation in the structure of herbaceous stems is shown in the cross-sections of spigelia (Plate 95); in ruellia (Plate 96); in the charts of powdered genuine horehound, powdered spurious horehound, and in the chart of powdered insect flower stems.
Cross-Section Spigelia Stem
Spigelia stem (Plate 95) has the following characteristic structure:
Epidermis
The epidermal cells are papillate.
Cortex
The cortical parenchyma cells consist of tan-gentially elongated cells which are oval in outline.
Phloem
The phloem consists of sieve cells, phloem parenchyma, and of bast fibres.
The sieve cells are small, and with thin, white, angled walls.
The phloem parenchyma resembles the sieve cells, but they are larger.
The bast fibres are rounded in outline and the walls are thick, white, non-porous, and non-striated.
Cambium
The cambium cells are rectangular in shape or the walls are collapsed and the cells indistinct.
Xylem
The xylem contains vessels, wood parenchyma, medullary rays. The vessels are small and angled, the walls are thick and white.
Wood Parenchyma
The cells are variable in size and shape, and the walls are thick. The medullary ray cells are small, narrow, and tangentially elongated.
Internal Phloem
External to the pith parenchyma are isolated groups of internal phloem consisting of sieve cells.
Pith Parenchyma
The pith parenchyma cells are oval in form and irregularly placed. The cells contain small, simple starch grains.
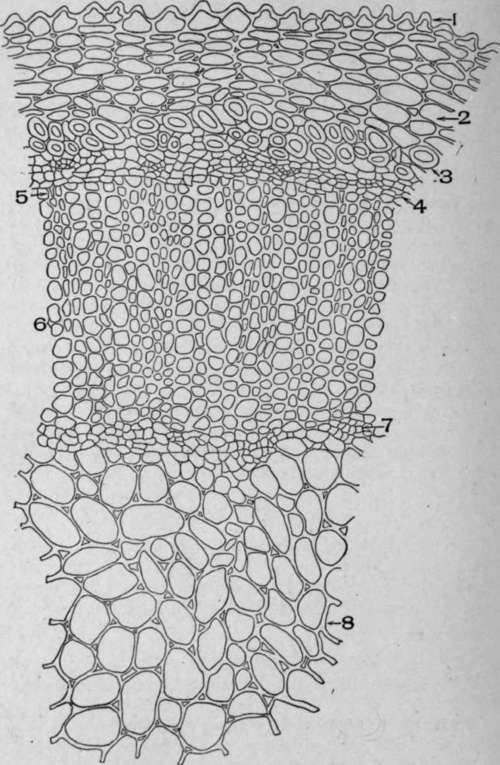
Plate 95. Cross-Section of Stem of Spigelia makylandica, L.
1. Papillate epidermis.
2. Cortical parenchyma.
3. Bast stereome.
4. Phloem.
5. Cambium.
6. Xylem.
7. Inner phloem.
8. Pith.
Ruellia Stem
The cross-section of ruellia stem (Plate 96) is as follows:
Epidermis
The epidermal cells are variable in shape and very large. There are no cell contents.
Cortex
The cortex consists of collenchyma and parenchyma cells and stone cells.
The collenchyma cells have very small, angled cavities and very thick walls. These cells make up the greater part of the cortex.
The cortical parenchyma cells are variable in size and shape. The stone cells occur singly or in groups. The walls are thick, white, porous, and striated, and the central cavity is frequently quite large.
Phloem
The phloem contains sieve cells, phloem parenchyma, and bast fibres.
The sieve cells have thin, white, angled walls.
The phloem parenchyma cells are frequently tangentially elongated, otherwise they resemble the sieve cells.
The bast fibres occur alone or in groups. The walls are thick, white and porous.
Cambium
The cambium cells are rectangular in shape and the walls are thin.
Xylem
The xylem contains vessels, wood parenchyma, and medullary rays.
The vessels are large; the walls are thick, white, and angled.
The wood parenchyma cells are variable in size and shape and the walls are angled.
The medullary ray cells are radially elongated and rectangular in shape.
Pith Parenchyma
The pith parenchyma cells are large and rounded in shape.

Plate 96. Cross-Section of Stem of Ruellia ciliosa, Pursh.
I. Epidermis. 2. Collenchyma. 3. Parenchyma. 4. Sclerids. 5. Bast fibres. 6. Phloem. 7. Cambium cells. 8. Xylem. 10. Pith parenchyma.
Powdered Horehound
The structure of powdered horehound is shown in Chart 97. The epidermal cells of the leaf (1) are wavy in outline, the guard cells are elliptical, the stoma lens-shaped, the epidermis often showing hairy outgrowth as in the illustration. The epidermal cells of the petals (2) have irregularly thickened beaded walls. The non-glandular hairs from the calyx (3); the long, thin-walled, multicellular non-glandular twisted hairs (4) from the leaves and stems; long, thin-walled, unicellular hairs (5) from the tube of the corolla; the glandular hairs (6) with a one-celled stalk and with two secreting cells divided by vertical walls; the eight-celled glandular hair (7) as seen in surface and side view; the spiral and reticulated conducting cells (8); the thick, White-walled fibres from the stem (9); the pollen grains (10) with nearly smooth walls.
The diagnostic elements of the U. S. P. horehound are the long, twisted, multicellular hairs (4), the glandular hairs (7), and the pollen grains (10).
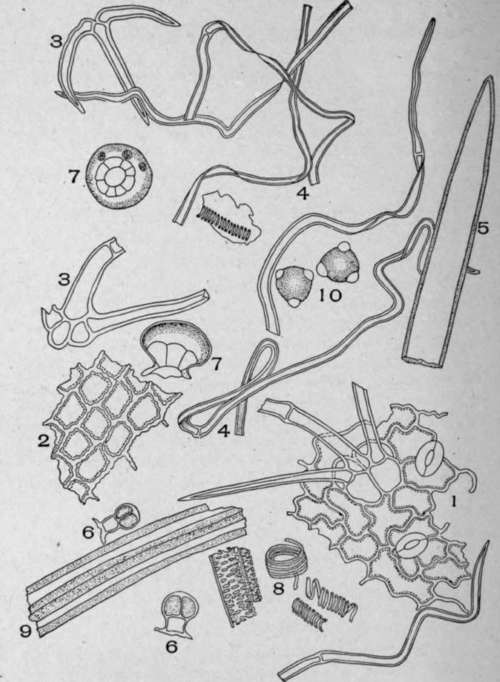
Plate 97. Powdered Horehound (Marrubium vulgare, L).
I. Epidermis of leaf showing the wavy epidermal cells, stoma, and a clustered hair. 2. Surface view of the petal epidermis. 3. Non-glandular hair from the calyx or corolla. 4. Long, thin-walled, twisted, non-glandular hairs from the leaves and stem. 5. Unicellular non-glandular hair from the tube of the corolla. 6. Glandular hairs with a one-celled stalk and with two secreting cells divided by vertical walls. 7. Surface and side view of the eight-celled glandular hairs. 8. Conducting cells. 9. Fibres from the stem. 10. Pollen grains.
Powdered Spurious Horehound
Marrubium perigrinum, which is a related species of horehound and which is a common adulterant of horehound, has the following structure (Plate 98):
The wavy leaf epidermis (1) with stoma; the beaded wall petal epidermis (2); the non-glandular, multicellular branched hairs (3) from the stem leaves or flowers; the broken pieces and branches of the compound hairs (4) scattered throughout the field; the glandular hairs (5) with a two-celled stalk; the eight-celled glandular hair (7) seen in surface view and a side view (8) of a similar hair; the long, pointed, unicellular non-glandular hair from the tube of the corolla, the wall irregularly thickened near the apex; the fibres (9) from the stem; the pollen grains (10) with prominent centrifugal projections; the conducting cells.
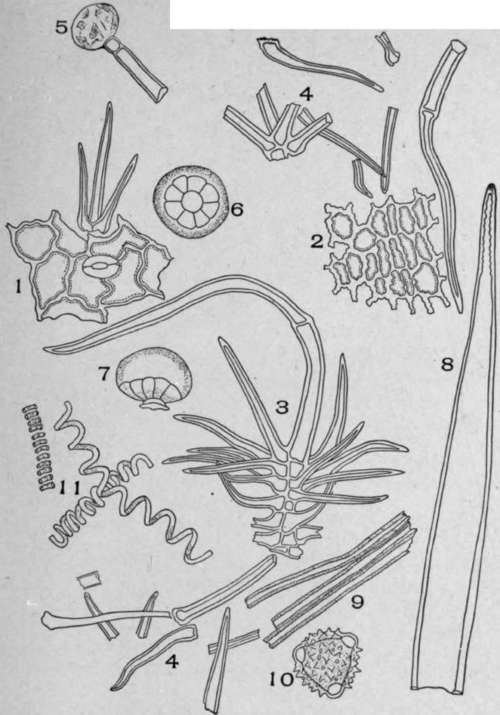
Plate 98. Spurious Horehound (Marrubium peregrinum, L.).
1. Surface view of the leaf epidermis. 2. View of the petal epidermis. 3. Non-glandular multicellular branched hair from the stem, leaves, or flowers with a few of the lower branches broken. 4. Broken pieces and branches from the compound hairs scattered throughout the field. 5. Unicellular glandular hair with a two-celled stalk. 6. Under-surface view of an eight-celled glandular hair. 7. Side view of eight-celled glandular hair. 8. Long, pointed, unicellular, non-glandular hair from the corolla, the wall irregularly thickened near the apex. 9. Fibres. 10. Pollen grains. II. Conducting cells of leaf.
The diagnostic elements of marrubium perigrinum are the multicellular branched hairs (3) which occur on all parts of the plant, usually much broken in the powder, with walls many times thicker than the walls of the hairs found in U. S. P. hore-hound; the pollen grains (10) with centrifugal projections and the stalked glandular hairs (5).
Insect Flower Stems
Insect flower stems are the chief adulterant of insect flowers. Until the passage of the insecticide law, it was a common practice to sell (for insect powder) a mixture of powdered stems and flowers. Since the passage of the law, the presence of the stems in a powder is supposed to be declared on the label. In spite of the penalties attached, their presence in a powder is frequently not declared, as evidenced by a microscopical examination of the insect powders obtained in the open market.
The structure of powdered insect flower stems (Chart 99) is as follows:
The epidermal cells of the stems are prominently marked with stoma and angled, striated wall cells (Fig. 1). On cross-section (Fig. 2) the stem is seen to be made up of epidermal cells with thick outer and thin side walls (Fig. 2). The T-shaped hairs (Fig. 3) are longer than those found on any other part of the plant. The fibres (Fig. 4) are the most characteristic part of the powder. They are elongated, and the walls are white and slightly porous and of nearly uniform thickness. They occur free in the field or in groups of two or more. The cross-section view of these fibres is shown in Fig. 5. The pith parenchyma (Fig. 6) is abundant and is composed of thick, porous-walled cells. On cross-section the cells are rounded and are separated by intercellular spaces. The conducting cells (Fig. 8) vary from spiral to reticulate.
![Powdered Insect Flower Stems (Chrysanthemum cinerariifolium, [Trev.], Vis.).](images/Powdered-Insect-Flower-Stems-Chrysanthemum-cinerariifolium.jpg)
Plate 99. Powdered Insect Flower Stems (Chrysanthemum cinerariifolium, [Trev.], Vis.).
1. Surface view of epidermis.
2. Cross-section of epidermis.
3. Hairs.
4. Fibres.
5. Cross-section of fibres.
6. Longitudinal view of pith parenchyma.
7. Cross-section of pith parenchyma.
8. Conducting cells.
Continue to:
My Books

