Plant
Description
This section is from "The American Cyclopaedia", by George Ripley And Charles A. Dana. Also available from Amazon: The New American Cyclopædia. 16 volumes complete..
Plant
Plant (Lat. planta), an organized being originating from a germ and nourished solely by inorganic substances. Although the differences between plants and animals in their higher forms are sufficiently obvious, with the lower ones it is difficult in many cases to decide whether an object ought to be classed as a plant or as an animal. Motion, which was formerly regarded as belonging only to animals, is now known to be a characteristic of some vegetable forms, and chemical constituents at one time supposed to belong only to plants have been found also in animals. Thus far it is not known that any animal can subsist directly upon the elements furnished by the mineral kingdom, while the plant is able to live upon air, water, and other inorganic materials, and assimilate them, converting them into organized structure, which may serve as food for animals; in doing this the plant takes in and decomposes carbonic acid, fixes carbon, and gives off oxygen, a function not known to belong to animals. Most animals have a stomach or internal cavity into which organized food is received; plants have no special receptacle for their food corresponding to a stomach. - Development of the Lower Plants. The plant in its most rudimentary form consists of a semi-fluid substance, protoplasm, surrounded by a cell wall - a minute sac or bag of protoplasm; in chemical constitution the cell wall is a carbohydrate, known as cellulose, its composition being carbon, oxygen, and hydrogen; the contents of the cell belong to the proteine compounds, and, besides the elements of cellulose, contain nitrogen.
An illustration of these simple forms of plant life is found in protococcus, which when present in large numbers appears as a green or even red scum, or in torula, the yeast plant. These plants, though so simple in structure and so insignificant in size, some of them not over 1/10,000 of an inch in diameter, while others are as large as 1/3500 of an inch, perform the two principal functions of larger and more complex plants: they grow and reproduce themselves. Being surrounded by a liquid (in the case of protococcus by rain water, which contains carbonic acid, ammonia, and other matters in solution), the little plant absorbs its nourishment through the cell wall; it increases in size by assimilating its food; small as it is, it can decompose carbonic acid, appropriating the carbon and setting free the oxygen, and combining this with the other elements form cellulose and protoplasm, which incorporated with that already in the plant increases its bulk to the full size proper to the species. When growth is attained, reproduction commences; the protoplasm subdivides into two, four, eight, or more masses, around each of which a cell wall is formed, and finally the segments separate, each portion growing to the full size and repeating the operation of subdivision.
Thus in a simple microscopic cell there is nutrition, growth, and reproduction. A step higher in organization are found plants of numerous simple cells placed end to end, as seen in some minute algae and fungi, in which a similar subdivision of the cells takes place, accompanied by a growth of the parts produced by division and elongating the chain of cells. In some of these microscopic algss the cooperation of two distinct cells is necessary to reproduction. A phenomenon called conjugation may be observed in some, such as zygnema, a very slender silk-like alga which forms a green scum of fine threads on pools in spring; a mioroscope of moderate power will show these plants to consist of elongated transparent cells placed end to end, within which the protoplasm is distinctly visible. The cells of two adjacent threads may be seen to bulge out toward one another; at length the projecting portions of the cells meet and finally touch, the cell wall between them is broken, and the contents of one cell pass over into the other, and mingling with the protoplasm of that cell form a spore, or reproductive body analogous to a seed, which being liberated starts life anew.
From this simple union of two different cells to form a reproductive body, it is but a step for the plant to have special cells devoted to reproduction, while the other cells of the plant are engaged in promoting its growth. The largest plant is made up of minute cells, each of which at some stage of its existence has passed through a life similar to that described for the pro-tococeus; it has been formed by the division of other cells, and has grown to its full size. - Germination and Growth from the Seed. A general view of plant life, as manifested in the higher forms of vegetation, may be best given by following a plant from the beginning. The first rudiment of the plant is to be found in the ovule or the forming seed. The ovule (the nature and structure of which will be explained further on) is the small pulpy body within the ovary which is to become the seed vessel, and which is itself to become the seed; a cell within the ovule is the beginning of the plant; this elongates by subdivision, and also increases in width until, taking the common bean as an example, two thick lobes are formed, connected by a short stem; when these parts are perfectly formed all growth ceases, and the seed is ripe.
If we soak a ripe bean in warm water until the skin can be removed, the parts may be seen, two lobes and a small stem uniting them, which in the bean is bent; this part of the seed (and in the bean it is the whole of it besides the seed coats) is the embryo. This is formed in every seed, though often much less distinctly than in the bean, and sometimes with only one lobe instead of two. The seed is to all appearance perfectly lifeless, dry, often very hard, and even bony, and it may remain dormant for years; some have been known to germinate after being kept in the ordinary way for half a century, and even after several hundred years when deeply buried in the soil; but the stories told of seeds taken from mummies several thousand years old and germinating have no foundation in fact. The essential conditions to germination are moisture, air, and heat; these must be present in proper proportions,-and are best applied through the medium of the soil, though germination will take place when these conditions are supplied in other ways. The first step is the absorption of moisture, which must be present in sufficient quantity, but for land plants there must not be an excess or the seed may rot; besides this, too much water excludes the air, without the oxygen of which germination is not possible.
It was formerly supposed that darkness was essential, but experiments have shown that germination is not influenced by the presence or absence of light. The temperature required varies with different seeds; while some will germinate at a few degrees above the melting point of ice, others, seeds of tropical plants, require 100° F. or more for their most successful germination; there is a degree of heat most favorable for each kind of seeds, though they will start at a much lower and will endure a much higher temperature; thus, while Indian corn will germinate at 48° and tolerate a heat of 115°, it is found that the process goes on most rapidly at the temperature of 93°. The time required for germination, even when the conditions are most favorable, varies greatly, and is of course greatly modified by temperature and other influences; some seeds sprout in 12 hours, those of most of the agricultural grains in four or five days, others require as many weeks, and there are several kinds which will lie dormant one and even two years. The time required by some seeds depends upon their treatment; if sown as soon as ripe they germinate the following spring, but if not sown until spring they remain dormant until the year following.
The growth of the young plant, which the embryo may be considered as soon as germination takes place, may be followed in the bean. The embryo when liberated from the seed coats consists of two very thick and fleshy leaves, a very short stem to which they are attached, and between them some rudimentary leaves, or a bud. The two fleshy leaves, commonly known as the seed leaves, are the cotyledons, the short stem is the radicle, and the small bud the plumule; the plumule is not at first visible in all germinating seeds, but it soon manifests itself. The first step in growth is the elongation of the radicle, the cells of which multiply and grow by a similar process to that by which it grew in forming the seed; if in the soil, one end pushes downward and the other makes its way upward, carrying with it the cotyledons, which it lifts above the surface of the soil. This growth is nourished by the material within the fleshy cotyledons; in the forming and ripening of the seed the cotyledons were stored to repletion with starch and other proximate principles; these are now rendered soluble and carried to the part where growth is going on, and are there assimilated by the growing cells, where some go to build up cell walls, and others to form cell contents.
These changes take place through the influence of diastase and other albuminoids, which act the part of ferments, converting starch from its fixed and insoluble condition into dextrine and sugar, forms which being soluble can be transferred. These changes are accompanied by the absorption of oxygen, the oxidation of some of the elements of the seed, and the escape of carbonic acid, and both hydrogen and nitrogen are also given off; and these changes are attended by the development of heat, which when germination takes place in a large quantity of seed, as in malting (see Brewing), becomes strikingly manifest. Before the store of nutriment in the cotyledons is exhausted preparation is made for drawing subsistence from the soil; roots, organs especially designed for absorbing liquids, make their appearance on the radicle, and through them water, holding various substances in solution, is conveyed to the interior of the plant, where this crude sap, as it is called, is converted into compounds capable of nourishing the plant.
Soon after the cotyledons of the bean appear above the soil the plumule increases in size; it is lifted up upon a stem, its rudimentary leaves enlarge and expand and disclose another bud, which in turn is raised by the growth of a stem and develops its leaves, and so the growth is continued. In the bean the early leaves are in pairs, but soon only one appears at a place on the stem, and the plant elongates by a successive addition of leaves, each separated from the one below it by a portion of stem; the cotyledons, though they become green like the other leaves, and no doubt to some extent perform the functions of foliage, after a while fall away. In examining other seeds and watching their germination, while the same general plan will be found to run through all, some marked deviations from the structure as seen in the bean will be noted. In the bean the seed leaves, having been diverted from their ordinary use and made the receptacles of nourishment for the young plant, are much distorted; a condition carried still further in the pea, in which they do not appear above ground; and in the acorn, horse chestnut, and others, they are so gorged that but little resemblance is to be traced between them and ordinary leaves; on the other hand, the cotyledons will often be found thin, and though differing in shape from the leaves produced later, they remain long after germination and are equally useful with the others.
In many seeds the nourishment for the growth of the young plant is placed outside of the embryo, and not within it, as in the bean; this material is termed albumen, and may be copious or scanty in proportion to the size of the embryo. The embryo is placed in the centre of the albumen, or at one end, or may be coiled in a more or less complete ring around the outside of the albumen. The embryo differs greatly in degree of development; in some cases the cotyledons are large and the plumule is distinctly visible, while in other seeds it is very rudimentary, the little stem or radicle having merely a notch at one end, the rude lobes thus formed foreshadowing the cotyledons. The name albumen is used to designate the nutritious matter placed for its nourishment outside of the embryo, without reference to its chemical composition; it was so called by the early botanists, who saw some analogy between its office and that of the white of an egg, and though endosperm, perisperm, and other distinctive names have been proposed, albumen is still used by the highest authorities. The character of the albumen varies greatly; it may be mucilaginous, fleshy, floury, oily, horny, etc. The presence or absence of albumen in seeds (albuminous and exalbuminous) is an important character in classifying plants.
A still more important one is found in the number of cotyledons, which in our example, the bean, is two, that number being found in the majority of the plants in northern climates, which are dicotyledonous. Plants in which but one cotyledon is present (monocotyledonous) are throughout their whole structure very different from the others, and this difference in the embryo, accompanied by other characters, serves for the separation of flowering plants into two great classes. In the conifers and some other plants the cotyledons often appear to be more than two, but these polycotyledonous embryos are regarded as dicotyledonous with the cotyledons subdivided. To return to the bean. Even in the very rudimentary state to which we have traced it, the bean just " up," as in the common expression, has all the organs of vegetation that it needs, all in kind that it ever will have, and indeed all that any plant has. Root, stem, and leaf, being all that are .concerned in the growth of the plant, its increase in size, are termed the organs of vegetation; the two last named already existed in the seed, while the root was formed as soon as germination was fairly accomplished.
The future growth of the plant consists in the repetition of these organs, and though in different plants these are wonderfully varied in form and size, and are sometimes turned aside from their proper functions, they may all be traced to these three elementary organs. In the plumule there is provision for the upward growth of the plant, though in a remarkable African genus, Welwitschia, no upward growth takes place, the long and leathery cotyledons being all the foliage that the plant, though supposed to live upward of a century, ever produces. In all plants the leaf or leaves are produced upon the stem at a point called the node; the space between one node and the next is the internode; the plant then, no matter how long it may grow, presents a succession of leaf-bearing nodes separated by internodes - in fact, is merely a repetition of the embryo, which was a very short stem or internode with a pair of leaves. In the plumule the undeveloped leaves were upon very short undeveloped internodes, and as growth went on these internodes elongated, and grew from nourishment afforded by the cotyledons at first, and afterward from that primarily taken up by the root.
The plumule is a very simple bud, but whether of this character or more highly organized, a bud (leaf bud) always consists of undeveloped leaves upon very short internodes; as the bean plant elongates there is still found at its upper end a simple bud, or what gardeners call the " growing point." - The Root. This may exist but a single season (annual), it may last for two seasons (biennial), or continue indefinitely (perennial). In a general way its functions are to absorb from the soil water and whatever it may hold in solution, and it also serves to fix the plant firmly to the earth. Its manner of growth will be mentioned further on; its chief office being that of absorption, it generally exposes the largest possible surface; in annual plants, where the work is to be done quickly, it is usually much subdivided and presents a mass of threads (fibrous), and these, and young roots generally, have their surface greatly increased by means of root hairs, which are prolongations like minute tubes, so small as to require a magnifier to see them, but which in the aggregate present an immense surface. If a main root descends into the soil, throwing off small branches from its sides, it is called the tap root; but its identity is often lost by its dividing into numerous small branches.
These branches may be thickened at intervals (a nodose root), as in the dropwort, or be entirely fleshy, as in the tuberous root of the dahlia. In some plants, especially biennials, the root, besides performing the usual functions, is converted into a storehouse for food; the roots of agriculture, the beet, carrot, turnip, and others, afford illustrations of this. The first year of these plants is passed in accumulating nutriment in the root, which becomes very much enlarged in diameter; but if such roots are examined while growing, they will be found abundantly supplied with rootlets or small branches, to carry on the proper work of absorption. If a hardy root of this kind, like the parsnip, be left in the ground over winter, or a tender one, like the beet, be set out the next spring, the growth from them is very rapid; a strong stalk is thrown up, flowers and seed are produced, and this mainly at the expense of the food contained in the fleshy root; and it is well known that when this second growth has fairly started the roots become exhausted of their nutriment and useless as food for man or beast.
Roots, whether one or several, which start from the lower end of the radicle, are primary roots; those which are produced elsewhere are secondary; many stems which trail along the ground (as squash and sweet potato) throw out secondary roots, and the propagation of plants by layering and from cuttings depends upon the ability of the stem to produce such roots. Some stems which do not touch the ground produce aerial roots; remarkable instances of this are found in the banian and the mangrove, and on a smaller scale in Indian corn, which for several joints above the soil throws out what farmers call brace roots, some of which reach the soil, while others do not; in the European ivy, the poison sumach, trumpet-creeper, and other climbers, the stem produces roots which are completely diverted from the usual office of the root and serve only to aid the plant in climbing, their main use being to attach them to the trees on which they grow, like the roots of epiphytes. - The Stem. The forms presented by the stem are still more numerous and varied; its proper function is to lift up the leaves to the influences of the light and air, and to keep up a communication between those organs and the root, as well as (generally) to bear the reproductive organs.
The differences in duration of the stem are similar to those of the root; the great variety in size, from the mere thread-like stem of some vines to the trunks 30 ft. or more in diameter of the giant trees of California and Australia, need only be alluded to. The direction of the stem, from the prostrate form of the cucumber, through the reclining stem of the wild currant, and the twining stem of the hop, up to the erect one of the forest tree, presents a great number of modifications, all of which in descriptive botany have their technical names. Perennial stems, such as trees, are usually terminated by a bud; this preparation to continue the growth of the stem is made during summer, and when such buds are examined they are found to contain rudimentary leaves crowded upon very short undeveloped inter-nodes; in the following spring growth from such buds is exceedingly rapid; the internodes elongate, while at the same time the minute leaves develop, and in many trees the whole growth of the season is made in a short time.
The terminal buds only prolong the main stems, but most stems branch, and the branches (as a rule) proceed from buds in the axils of the leaves, or where these join the stem; as leaves are opposite or alternate, the branches are thrown off in a similar manner; but as only a small portion of the axillary buds develop, the arrangement of the branches is not so symmetrical as might be expected. In trees and shrubs the buds, both terminal and axillary, are prepared beforehand and remain dormant through the winter. In many cases the development is continuous, especially in annual stems; buds are developed as soon as formed, and we find upon such stems every gradation between the fully developed internodes, through gradually shorter ones, to a loose bud or growing point. Buds which are neither terminal nor axillary, but which appear anywhere upon the stem, are termed adventitious; some trees and shrubs produce them habitually, while others do so only when the regular buds have been destroyed. To some forms of the stem special names are given, some of which are in common use; a weak prostrate stem, like that of the strawberry, is a runner; a sucker is a branch starting upon the stem, at a point below the surface of the soil; a stolon is a stem which bends over and takes root at the end; these are all taken advantage of in propagating plants.
In some cases a branch is developed as a tendril to aid the plant in climbing; there are tendrils of a different nature, but a fine example of this form is afforded by the beautiful Virginia creeper, in which a slender branch has at the end of each subdivision a disk which attaches itself to a wall, after which the branch coils into a spiral. Thorns or spines are often suppressed branches, which become indurated and sharp at the point, as maybe seen in a wild pear tree, the thorns, and many of the shrubs belonging to the arid western plains, where almost all vegetation seems to be on the defensive. In most of the cactus family the leaves are mere rudiments which soon fall away, and the thick, green, fleshy rind of the stem performs the functions of both leaf and stem. - These are a few of the above-ground forms of the stem, but those found below the surface are nearly as numerous and varied; much of that portion of the plant which, being found below ground, is popularly considered as the root, really consists of branches belonging to the stem.
As there are creeping stems, which lie upon the surface and throw out roots at each joint or node, so there are plants with similar stems which behave in the same manner just below the surface; these stems have nodes, from which roots proceed; but having no use for leaves, these organs are represented by mere rudiments in the form of scales; they have terminal and axillary buds, and if separated from the plant are capable of living, and if they are cut into as many pieces as there are joints, each node is able to make a plant; some weeds, eminently couch grass, and some sedges in sandy soils, have underground stems of this kind, which ordinary cultivation only breaks up and subdivides, and thus multiplies rather than destroys the weed. Such an underground stem is called a rhizome or root-stock, which may be slender as in the couch grass, or very much thickened and fleshy as in the iris, bloodroot, and many others; in some the growth is very slow and definite, as in the Solomon's seal of our woods (smila-cina), which throws up its leafy stem, and during the season matures a bud at its extremity; when the stem dies it leaves a scar on the rootstock; the bud thus formed will produce an above-ground stem the next year, another bud will be formed in advance of it, and so on, the plant gradually moving forward in this manner, while the older part of the rootstock, being no longer of use, slowly decays.
Another form of the rootstock is the tuber, as the potato, which is only a very fleshy rootstock, gorged with starch to serve as nutriment for the numerous axillary buds (eyes) which it bears. In the Jerusalem artichoke the relationship between the tuber and the proper stem is more distinctly seen; it appears at the end of a much shorter stem than the potato tuber, and the scales which stand for leaves are much more prominent. Another form of underground stem is furnished in the corm, which is often called a bulb; and it is a solid bulb, or very short, more or less globular root-stock, of which among garden plants the gladiolus and crocus, and among wild plants the Indian turnip or Jack-in-the-pulpit (arisama), are familiar illustrations; that these are proper stems, though much reduced, is shown by the leaf scars and axillary buds they bear; and that they are not proper roots is shown by their bearing distinct roots. The stem of the cyclamen is just like a corm, but grows above ground. The scaly bulb differs from the corm or solid bulb in being clothed with fleshy scales, which are the bases of the leaves, fleshy and thick with the nutriment stored in them by the upper portions of the leaves, which have fallen away.
A longitudinal section of a scaly bulb, like that of a lily, shows the stem reduced to a mere plate, to the upper surface of which the scales are attached, and in the axils of these are a greater or less number of axillary buds; from the lower part of this reduced stem the roots are produced. The similarity between the underground stem of the scaly bulb, with its shortened inter-nodes and fleshy scales, and that of the similarly shortened above-ground stem of a house-leek or an echeveria, bearing short fleshy leaves, is very striking. All bulbs are not scaly, some, as in the onion, having the bases of the leaves in concentric layers. - The Leaf. This, the remaining organ of vegetation, is described as to its various forms and its functions under Leaf. In most of the higher orders of plants the root absorbs nutriment from the soil, and the leaves digest it and prepare it for increasing the growth of the plant, and the stem serves as a medium of communication between the root and the leaves. Some of the many shapes assumed by the root and the stem are here pointed out, and those of the leaf are given elsewhere, at least a sufficient number to show that the wonderful variety presented by plants may be produced by very simple modifications of the root, stem, and leaf; and it has been suggested (of which fact careful observers will find numerous other illustrations) that these organs may perforin more than one office, as the leaf may serve both as leaf and tendril, and that they may serve a purpose widely different from their ordinary one, as when roots aid the plant in climbing. - Internal Structure. The organs of vegetation have been treated only as to their external forms, except the leaf, which has its internal structure described in another article. (See Leaf.) When the embryo is fertilized a new cell is formed; the future growth of the embryo is due to the subdivision and multiplication of that cell; and when growth is resumed in germination, the future plant results from the multiplication of the cells contained in the embryo.
The microscope shows the plant to be made up in all its parts of cells, which vary greatly in size and form, but are all regarded as modifications of the simple cell. The cell when not pressed upon by others is of a more or less spherical form, such as has been described in the simple cellular plants; it is sometimes marked by a lining which, being thin in places, gives a dotted appearance; it is sometimes nearly filled with a deposit, as in the gritty cells so common in the fruit of the pear. An aggregation of simple cells forms cellular tissue, like that found in the pulpy portions of the leaf, in young stems, in fruits, and in other soft parts of ,the plant; a cross section of the young stem shows the cellular tissue to be polyhedral, and the cells have their longest diameter in the direction of the growth. The walls exhibited by a cross section of cellular tissue are structurally double, and may often be separated; and frequently, where the union of contiguous cells is not complete, intercellular spaces are left. The size of the simple cell varies greatly; in the fruit of the orange family it is from one fourth to one half of an inch long, but they are ordinarily much smaller, varying from 1/1200 to1/200 of an inch.
The woody part of a plant shows cells of a different form; they are many times longer than wide, and are placed lengthwise parallel with the axis of the stem; their smallest diameter is from 1/1000 to1/2000 of an inch, but in some cases much larger; they are tapering at the ends, and overlap one another in such a manner as to make the woody tissue they compose very strong; the term fibro-vascular is also used for this tissue. A cross section of woody tissue shows that their walls are thick and tough, though with numerous thin spaces; in old or heart wood, the cells are nearly filled with a woody incrustation; the wood cells or woody fibres in the pine family are remarkably large, and are marked by conspicuous dots, which are thin spaces in the cell wall; these are so characteristic that the wood of the pine family, even in the fossil state, may be at once recognized by the microscope. The cells of the inner bark are usually longer, finer, and tougher than those of ordinary wood, and are termed bast cells or bast fibres. The bast tissue is so developed in the linden or basswood tree that it is used for coarse fabrics; it is this that constitutes the usual fibres of the flax and hemp plants, and of others which furnish fibre in their bark.
Mixed with the woody fibres, and modifications of them, are ducts or vessels, as they are sometimes called; they are much larger than the ordinary wood cells, and are often continuous tubes of considerable length, evidently formed from a row of cells placed end to end with the intervening partition obliterated; these are dotted by the appearance of thin places when young, and of pores when old; other ducts are found with the lining disposed in rings (annular ducts), and others (spiral vessels) are lined by a continuous spiral thread, which may be seen on carefully breaking a rose leaf across and gently separating the two parts. Besides these, there are milk tubes and oil tubes in plants, with special secretions, and intercellular spaces. The cell is the elementary organ of the plant, every portion of which is made up of an aggregation of cells of some kind. In the germinating seedling the stem or radicle is mainly cellular tissue, but very early there appear bundles of woody fibre, which increase in number as the stem grows upward and produces leaves. - Growth of Dicotyledonous Stems. In dicotyledonous plants the fibro-vascular bundles are placed to form a ring between the centre and circumference of the stem; in annual stems the woody fibre is not so strong or so abundant as in perennial stems; in the former there is often a considerable space between the bundles of wood, while in the other they are crowded and show on a-cross section as a nearly complete ring separating the central portion or pith from the outer portion or bark; it will be noticed that plates of cellular tissue pass from the central pith between the bundles or wedges of wood and keep up a communication with the outer portion, or bark.
A cross section of a woody stem of a dicotyledon shows in the centre the pith, then a ring of wood, and outside of this the bark, with medullary rays or lines of pith running through the wood and connecting the two; the pith is simple cellular tissue; the wood, besides proper woody fibre, contains spiral vessels and ducts, often so large as to be visible without a glass. The bark, which early in the life of the stem was simply cellular tissue covered by the epidermis, is found at the end of the season to consist of several parts; that portion next to the wood of the stem has bast cells formed in it, and becomes the inner bark or liber; outside of this is the green layer, which is much like the cellular part of leaves; this soon becomes covered by the corky layer, and to this is due the color most woody stems present at the end of the season; in one oak this layer becomes unusually developed, and is separated as cork, for which no substitute has as yet been found. Outside of the whole is the epidermis, a layer of thick-walled flattened cells.
In a stem of this kind, when growth commences the second spring, the bark is readily separable from the wood, and between the two will be found a mucilaginous substance called the cambium; this consists of extremely delicate forming cells, as it is here that the growth of the stem takes place; the inner portion of the cambium belongs to the wood and the outer to the bark, and in this a layer of new wood forms outside of that of the previous year, while a layer of liber or inner bark is added inside of that of the year before, the medullary rays also elongating so that the wood of the second year is also traversed by these. The increase in the woody system of the stems of dicotyledons taking place always by additions on the outside of the old wood, such stems are exogenous, or outside-growing, and the term exo-gen is used as synonymous with dicotyledon, even when the plant is only an annual. In exogenous stems this growth goes on year after year, a layer of wood being annually added, and in the trees of temperate climates the number of these rings can be readily counted; the woody fibres first formed in the spring are larger and less dense than those formed later when vegetation is more sluggish, and this difference in texture ajlows the line of demarcation to be seen, when there are not, as is sometimes the case, ducts in the wood first formed.
In most old stems the wood near the centre after a while becomes solidified by the incrustation of the cells, forming heart wood, which is essentially dead, as it takes no part in the growth of the tree; the newer wood in which vegetation is still active is popularly known as sap wood. While the bark receives an annual layer upon its inner surface, the outer surface undergoes marked changes; it is acted upon by the elements, and soon loses its epidermis, often the corky layer, and eventually the only part left is the liber, which gradually dies on the exterior, and is renewed on the interior; in some trees the bark falls away in flakes, in others in strips, the method of getting rid of the old bark varying greatly in different species. In a very old exogenous tree the only living portions are the buds which continue the growth and from which foliage is developed, the fresh portions of the roots, and the new layers of wood and bark, with the intervening cambium which connects the roots with the buds; examples of hollow trees are not rare in which all the remaining portion of the stem has been removed by decay.
The internal structure of the root is similar to that of the stem; its chief increase is by lengthening, and this takes place much more rapidly at the tip than elsewhere, or rather a very short distance back of the tip, the very point being a sort of cap or cushion of dead cells which serves to protect the living point as it pushes its way through the soil; in some plants this root cap is very distinct. It was formerly supposed that the root was furnished with special absorbing organs, which were called spongi-oles, a term still sometimes used, though it was long ago shown that such organs have no existence, and that the root absorbs anywhere through its recently formed parts. The root differs from the stem in branching irregularly and in usually bearing no buds, though under certain conditions it will, like other parts of the plant, form adventitious buds; and many plants difficult to multiply otherwise are propagated by root cuttings, which when separated from the main plant will produce buds. - Mo-nocotyledonous Stems. In the stem of monocotyledons a cross section shows no distinct ring of wood, and no separation into pith, wood, and bark, but bundles of woody fibre are irregularly scattered all through the cellular tissue or pith, though less numerous in the centre than at the circumference of the stem; these bundles have their origin in the leaves, and as new leaves are developed new bundles push their way down through the central portion of the stem, in young plants reaching the roots, but in older ones finally curving outward and losing themselves in the rind.
In these stems the newly formed wood is deposited within the older bundles, and they are called endogenous, or inside-growing, endo-gen being synonymous with monocotyledon. The internal structure of a stem of this kind is shown in the article Palm; and the corn stalk illustrates that of an annual endogenous stem, which when young has an epidermis, but no separable bark. Each one of its fibrous bundles contains all the elements of the exogenous stem; a microscopic examination of such a bundle shows bast cells, wood cells, ducts of various kinds, and a portion in which all the growth of the bundle is carried on, corresponding to the cambium in dicotyledonous stems. - Epidermal Tissue. The epidermis has been mentioned as clothing stems of both kinds when young, and indeed it covers all parts of the plant exposed to the air, save the stigma, in which the naked cellular tissue is exposed, but usually covered by a viscid secretion; the general structure of the epidermis is illustrated under Leaf, where it is shown to be a layer of flattened empty cells with numerous openings (stomatce), or breathing pores; in some cases this has a deposit of silex and is very hard; in others it is covered with a waxy secretion, as in the bloom of the grape and other fruit, and in the leaves of the cabbage.
In some cases the exposed cell wall is developed as a cuticle above the proper epidermis. The cells of the epidermis are sometimes in flowers prolonged into papillaa, which give a peculiar velvety appearance; again they are produced as simple hairs, or several cells may unite to form a hair, which may be still more complicated as a sting to exude an irritating fluid; glands and prickles often belong to the epidermis. Upon the root the epidermis is very thin, and is not upon the tender growing point, but is formed later by the thickening of the exterior cells. - Physiology of Plants. In the simple plants mentioned at the beginning, one cell performs all the functions of growth and reproduction; whatever part of a higher plant be examined, the cell in some form will be found, and growth wherever it takes place in the plant consists in the multiplying of cells. The cell, whether in the soft pith, the hard wood, or the remarkably strong tissue of the inner bark, without reference to its contents, is chemically the same. The ultimate composition of the cell itself is carbon, hydrogen, and oxygen (C12H20 O10), and is called cellulose; it has the same composition in all plants.
For the formation of cellulose these elements must be supplied to the plant; they are not converted directly into cellulose, though this is their ultimate destination, but into dextrine, sugar, gum, and similar principles, either identical in composition with cellulose, or differing from it only in having a few equivalents of oxygen and hydrogen more, or cellulose with the addition of water; all these principles, including cellulose, being essentially carbon and water, are termed carbohydrates. Water is supplied by the soil; carbon is furnished in the form of carbonic acid (C02), which is constantly present in the atmosphere in the gaseous form, and in the water of the soil in solution. The decomposition of carbonic acid, setting free its oxygen and combining its carbon with the elements of water, is the striking phenomenon attendant upon plant growth. This change is effected only through the agency of chlorophyl (leaf green), and in the leaves or those parts capable of performing the functions of the leaf; as its name implies, chlorophyl is usually green, but it is sometimes of another color.
Giving the green color to leaves, one would suppose it to be present in very large quantities, but an examination of the cells of a leaf by the microscope shows the proportion to be surprisingly small; it is seen in the form of exceedingly minute grains, attached to the walls of the cell or distributed through its contents. It has been stated that the foundation of the cell is protoplasm, and that the cell wall is built over that; protoplasm, also called the formative layer, is found in all cells of the growing parts of the plant; and chlorophyl grains are formed in it and belong in it. Protoplasm, so essential to the growth of the cell, differs from it in chemical composition, as it contains nitrogen in addition to the elements before mentioned. Other vegetable principles have a similar composition, often with the addition of sulphur and phosphorus, and are called, from their resemblance to albumen, albuminoids. To form these the plant must be supplied with nitrogen, and although this is present in the atmosphere in such abundance, the most careful experiments fail to show that the plant appropriates it. It enters the plant in combination as nitric acid in the form of nitrates and as ammonia, these being in solution in the soil.
These elements, carbon, oxygen, hydrogen, and nitrogen,, when the plant is burned or decays, entering into new combinations, disappear in the gaseous form, and hence are termed the volatile elements; but there is left behind the ash, or non-volatile portion, the source of which must have been the soil. It was formerly supposed that the presence of non-volatile or earthy matters in plants was accidental; being dissolved in the water of the soil, they were carried into the plant and there deposited. "While more of these than is useful may be taken up, and some are of no known use, it is now well established that others are absolutely necessary; it is sufficient to say here that potash, lime, iron, phosphoric acid, and some others play an important part in vegetation, promoting the activity of growth, aiding in the diffusion of albuminoids, and in other ways; it is also ascertained that chlorophyl, upon which so much of the work of the plant depends, is not formed in the protoplasm unless iron be present, though an infinitesimally small quantity is required. These are the essential constituents of plants, furnished in part by the atmosphere, and in part by the soil.
The volatile elements of plants may be regarded as coming primarily from the atmosphere, though in good part immediately obtained from the decay of former vegetation; and the vegetable products which cover the earth in the living state, or partially decayed in the soil as humus, or elsewhere as coal, peat, and muck, together represent the amount of materials that plants have taken from the atmosphere. - The conversion of the inorganic, lifeless elements into organized matter and structure endowed with life (assimilation) is done in the leaf, and a reference to its structure (see Leaf) shows how well it is adapted to the work; it is so formed as to present the greatest amount of surface to the sun and air, and in its interior structure it is mostly loose cellular tissue with abundant air spaces, which communicate with the external air through innumerable openings (stomata) in the otherwise impervious epidermis. A similar structure is found in other parts which act the same as leaves; in many cacti and euphorbias the whole leaf work is done by the green stem, and the young stems of other plants do the same, as do the calyx and some other parts homologous with leaves, such as the developing fruit.
The roots absorb from the soil water containing in solution the ash ingredients or mineral substances, nitrogenous and other matters, which (without here noticing the means) is carried into the plant and ultimately to the leaves; this crude sap, as it is called, does not, as many suppose, move upward in distinct currents through regular channels directly to the leaves to be elaborated, and afterward descend by other channels to contribute to the growth of the plant. There is however an abundant motion of the juices of the plant, and this takes place in any direction required. As soon as the liquid taken up by the soil enters the plant, it meets with elaborated sap in the cells, and in all parts of the plant is a complex liquid. The leaves gather carbonic acid gas from the air; this is decomposed within them, and an equivalent quantity of oxygen is returned to the atmosphere; this decomposition of carbonic acid, which in the laboratory can be done only with difficulty, is effected in the delicate tissues of the leaf, but only in direct sunlight; from the carbon thus obtained and the elements of water carbohydrates are formed; this process takes place apparently within the chlorophyl cells, and in some manner requires the aid of the ash ingredients of the sap.
To form albuminoids the nitrates and other nitrogenous compounds taken up from the soil furnish the needed additional element. The successive steps in assimilation are not known with certainty, as this wonderful chemistry is carried on within the recesses of the leaf, but the general fact of the decomposition of carbonic acid is well established by experiment. The interesting relations between plants and animals cannot be discussed, but it may be stated in passing that all the food of animals is provided directly or indirectly by plants, and that in preparing food for animals plants take from the air carbonic acid, destructive to animal life, and return to it oxygen, essential to the respiration of all animals; and that animals on their part return a large portion of the plants they consume as food to the air in the form of carbonic acid; thus these two great divisions of the organic world, plants and animals, reciprocally act upon the atmosphere, each contributing to the welfare and supplying the wants of the other. The form in which assimilated matter first appears within the plant is probably that of dextrine and glucose or grape sugar, principles readily changed into other soluble carbohydrates or into the insoluble form of cellulose.
Another important carbohydrate is starch, like cellulose insoluble, and the form in which assimilated material is kept in reserve; it is readily converted into any of the soluble carbohydrates, and ultimately into cellulose; hence we find it wherever nutriment is provided for future growth; it constitutes in some form a large part of most seeds, and is stored near buds, notably in tubers and fleshy roots, to supply the demands of the growth of young shoots. The growth of the plant from reserve material has been already noted in describing germination; the phenomenon is shown in a striking manner in the potato in spring, which will often make shoots several feet long in a dark cellar, the cells of these shoots being cellulose formed from the starch of the tuber; but before the starch can contribute to this growth it must be rendered soluble, that it may be moved to the part where growth is going on; it is well known that a potato in this condition is sweet and " soggy," and unpleasant to use as food, a condition due to the conversion of its starch into glucose (grape sugar), dextrine, and other soluble forms.
The change of one form of carbohydrate into another is quite distinct from assimilation, and the name metastasis has been given to it, as the equivalent of the German Stoffwechsel. It is unnecessary here to consider other principles found within plants, most of which are treated in separate articles, such as fats, oils, resin, caoutchouc, special principles like quinine and morphine, acids, and many others which, though of great importance to man as vegetable products, do not, with some exceptions, take part in the growth of the plant, and often result from a conversion of some of the other principles. - Movement of Liquids. The motion of liquids within the plant is quite complex. Within many cells, and probably in all, when young, the protoplasm is in motion; in some cases the whole body of it is seen to rotate very slowly, though under the microscope, the motion being magnified, it appears rapid; Val-lisneria, the tape grass or eel grass of fresh water, affords a beautiful microscopic object on account of this rotation; in other cases the motion is confined to portions of the protoplasm, and is seen in streams or strings running in various directions; this movement in one cell is quite independent of that in adjoining cells, and has no relation to the passage of liquid from one cell to another; it is regarded as a vital movement of the protoplasm.
The motion of liquids from place to place within the plant, as from the root upward, is due to several causes. In the process of assimilation large quantities of water are evaporated; a great deal of water is needed as the vehicle by which nutriment is brought from the soil, and to hold the assimilated material in solution, and exhalation of watery vapor goes on rapidly in growing plants. This vapor is mainly exhaled through the stomata or leaf openings, and its amount is governed by the hygrometric condition of the atmosphere; in a damp day but little transpires; in a long continued drought evaporation might continue greatly to the detriment of the plant were not the stomata so constructed (see Leaf) as to shut off nearly all communication with the interior of the leaf. The common agricultural plants during five months of growth exhale 200 times their dry weight of water. The tissues of the plant being gorged with liquid, when evaporation takes place in the leaves there is a demand upon the parts below to supply this loss, and thus an upward flow is established from the roots toward the leaves, and the amount of water taken up by the root is in direct ratio to that given off by the leaves. If the demand made by the leaves is greater than the root can supply, the plant droops.
It was formerly thought that the leaves absorbed water from the atmosphere, but experiment has shown this to be erroneous. Though the cells and ducts differ greatly in length, the tissues of the plant are really made up of closed cavities, the walls of which, though no openings have been detected, are permeable by liquids, and the movement of sap within the plant consists of a transference from one cell to another. Several physical forces are probably concerned in the movement, specially that known as osmose. When two liquids of different densities are separated by a permeable membrane there is a tendency to interchange, a small quantity of the denser liquid passing into the lighter, and a much larger quantity of the lighter liquid passing through the membrane into the heavier; this force, called osmose, explained at length in works upon physics, and easily demonstrated (see Endosmose), is sufficient to account for much of the sap movement and transference of the fluids from cell to cell; if the contents of the leaf cells are made denser by evaporation, the less dense liquids of the cells near by will flow toward them, and the movement be communicated from cell to cell down to the roots.
Some of the phenomena of the movement of liquids are attributed to capillary or surface attraction, and the power of membrane to separate solutions of different kinds, discovered by Graham (see Dialysis), is evidently concerned in the nutrition of the plant. While there are no regular ascending and descending currents of sap, as was formerly supposed, the liquids in the different tissues of highly organized stems are not all alike; the ascent of water or crude sap from the root is mainly through the newer wood, and the movement of formative material or elaborated sap to contribute to the growth of parts below the leaves is through the young bast or inner bark. But growth is taking place in other parts of the plant, new stems and leaves are forming beyond the assimilating leaves, and in most cereal grains the growth of the fruit is quite above all the foliage and consequently not to be reached by descending sap. The motion of the assimilated material is toward all points in which growth is taking place, or to those in which it is to be stored as a reserve; and whenever it is appropriated, either by forming new cells or stored up as starch, these points become centres of attraction, and the movement of the material is in their direction. - Automatic Movements and Irritability. The movements of plants may often be readily observed; many leaves take on a different position at nightfall, as may be seen in the common locust and wood sorrel (see Oxalis), and many flowers which close at night or in dull days; in the evening primrose (see CEnothera) the petals open with a sudden jerk.
The "movements of climbing plants" are admirably described by Darwin in a memoir with that title; the free end of a twining stem makes a rotary movement with a rapidity differing with the species, the hop on a warm day describing a circle in one to two hours; when the stem comes in contact with a support, it winds around it; the phenomenon can only be observed when such stems extend above the support and are free to move, in which case the upper internodes are seen to revolve as if in search of something to twine upon. Tendrils move in a similar manner; when the hooked end of a tendril catches a twig or some other object, the tendril twists into a coil, as if by shortening itself to draw the plant nearer to the support, one half of the coiling being in an opposite direction to the other half; if the tendril fails to catch hold of some object, it coils after a while, but in this case the spiral runs all in one direction. Tendrils are irritable, and coiling may be induced by slight friction. These voluntary or rather automatic movements are very slow when compared with those of some of the lower algse, which like oscillaria are constantly writhing.
In the higher plants this motion is seen in the flowers of some orchids, in which the lower petal or lip is raised and lowered with a regular movement; desmodium gyrans, sometimes seen in greenhouses, has trifoliate leaves, of which the two lateral are constantly in motion, moving by a series of visible jerks day and night. These movements are independent of any external agency, while others are due to irritability, as may be conspicuously seen in the sensitive plant (see Mimosa) and some others; the irritability of tendrils is above referred to; another striking instance of this property, apparently connected with something corresponding to a nervous system, is presented by the Venus's fly-trap. (See DionAEa.) Nor is this irritability confined to leaves; an Australian orchid shuts up its lip when an insect alights upon it and encloses the captive, and the stamens of the common barberry move if touched. - Propagation of Plan ts by Division. In m any plants there is ample provision for multiplication of the individual by means of subdivision.
The strawberry strikes root at each joint of its prostrate stem, a bud is formed there, and at length the stem dies, severing the connection with the old plant, and leaving these rooted portions to shift for themselves as new plants; the same thing happens with other above-ground stems, some of which only take root at the tip, which bends over and reaches the ground apparently for this very purpose. Propagation by means of underground stems or rootstocks is still more common; the rootstocks of the tall blackberry, the passion flower, and others, run along below the surface for several feet, and throw up a shoot at the end, which in time becomes an independent plant. As may be seen in some lilies, in dioscorcea, and other plants, buds which have been formed in the axils of the leaves fall to the ground and there take root to form new plants. These methods of multiplication are extensively imitated by the cultivator, and the operations of layering and dividing plants are in imitation of nature's methods; and we carry it still further in propagating by cuttings, which is founded upon the fact that the stem, and in many plants even the leaf, will under favorable circumstances form adventitious roots; the majority of plants if treated at the proper time may be multiplied in this way, in many each node being capable of forming a new plant.
In grafting and budding the cutting, instead of being planted in the soil, is set in the tissues of another but related plant. All such propagation, whether it takes place naturally or is done by the aid of man, merely subdivides the individual, and each new plant thus obtained possesses all the peculiarities of the parent. Thus all the potatoes of any given variety, the Early Rose for instance, of which there are untold millions raised each year, are subdivisions of the original plant; and so the hundreds of thousands of Bartlett pears, all over the world, are in reality parts of the original seedling. That these subdivisions practically retain the peculiarities of the parent plant without variation, except the slight modifications caused by a more or less congenial soil, and manifested in size and vigor, is a fact of the highest horticultural importance. - Inflorescence. Whatever may be the provision for this kind of multiplication in nature, sooner or later the plant multiplies in another manner, by seed. In mentioning the multiplication of the lower orders a plant was cited in which the contents of any two of its cells unite to form a reproductive body or spore.
In plants of a higher organization the reproductive body, the seed, is formed by a distinct set of organs, the organs of fructification, and these are collectively the flower. In annual plants the production of flowers often commences when the plant is very young, only a few weeks from the seed, while on the other hand perennial plants, especially trees, may go on repeating their organs of vegetation, or growing, for many years; some, like the century plant, which in its native country blooms in 10 or 15 years, spend all their lives in preparing for the event, and when they have bloomed die, as if exhausted with the effort; while others continue to bloom, if not yearly, at intervals for centuries. Flowers appear either at the end of the stem or in the axils of the leaves, just where leaf buds appear, and like them are either terminal or axillary. Axillary flowers appear at the base of ordinary leaves, usually upon a little stalk or pedicel, but more frequently the leaf is reduced in size; these smaller leaves are termed bracts; if at the same time the internodes of the stem are shortened, one of the commonest forms of flower cluster, the raceme, is produced; if the flowers are without pedicels, or sessile, and crowded, a spike is the result; and the pedicels of a raceme may be branched and each branch bear a flower, producing a panicle.
These forms of inflorescence are readily seen to be analogous to ordinary branching; others, like the corymb, umbel, etc, appear to be more complex, but are only modifications of the forms already noted. In a corymb the internodes are short, and the lower pedicels so elongated as to bring their flowers on a plane with the upper ones; in the umbel the internodes are obliterated, the pedicels (rays) all starting apparently at the same point, and of equal length, while the bracts are brought together in a whorl to form an involucre. In the raceme the lower flowers are the oldest; and in many cases the stem or common axis keeps on elongating, so that above new buds are developing while the flowers below have fallen, and is said to be indeterminate, though generally the growth of the stem or axis sooner or later ceases. In the umbel the outer flowers represent the lower ones of the raceme, and are consequently the oldest; the flowering, beginning at the circumference and proceeding toward the centre, is centripetal, a term applied also to the raceme and similar forms.
Where the flower is at the end of the stem, that ceases to elongate, and such inflorescence is determinate; flowers of this kind are often solitary, and when they form a cluster it is by the production of other flowers upon the stem below the terminal one. In such inflorescence a flat cluster, the cyme, corresponding to a corymb, may be formed, but the oldest flowers will be in the centre; such inflorescence is centrifugal. The term ament or catkin is applied to an elongated cluster of scale-like flowers; a spadix is a fleshy spike upon which the flowers, often imperfect, are closely crowded, as in the Indian turnip, the palms, and others; this is often protected by a leafy covering, the spathe which in the Indian turnip, and especially the calla, becomes showy and commonly passes for the flower. There are other forms of inflorescence which have distinctive names, but they are all traceable to the simple forms here noted. - Structure of the Flower. The flower has for its end the production of seeds, and all its parts are subservient to that. Numerous as are the forms presented by flowers on every hand, they may all be referred to one simple plan.
Instead of taking the simplest form of the flower for illustration, the plan of its structure may be best understood by the examination of one of the more complete forms, one which has all the parts to be found in any flower. There are many flowers which might serve as a pattern or type, but there are reasons for using the one selected by Gray in his admirable " Lessons," one of the sedums or stonecrops. An examination of this flower shows, beginning from without, a series of five green, leafy parts, which together form the calyx; the parts are sepals. Within these are five more delicate (and in this case white) leafy bodies, called petals, which form .the corolla; and these two together are the floral envelopes. Immediately next to the corolla, toward the centre of the flower, is a series of (in this case 10) bodies, the stamens, quite unlike calyx and corolla, being slender threads with a knob at the apex of each; and quite within all the other three series is a cluster of five green parts, the pistils. As the two outer series are termed the floral envelopes, the inner two, the stamens and pistils, are the essential organs. All of these, in the order here named, are placed upon the end of the flower stalk, which often has an enlargement to hold them, called the receptacle.
A longitudinal section through the flower would show each series placed one above the other upon the receptacle; or their position may be made plainer by an illustration giving two of each series detached, but in their relative positions. The calyx is generally green and leaf-like in texture, though in some plants, as the fuchsia, it is colored, a term used in botany for any other color than green. The corolla is of a more delicate texture and is very rarely green, but is usually the most showy part of the flower. The stamens have two distinct parts, the stalk or filament, and the anther or knob at the top; the anther is the important part of the stamen, as the filament may be short or wanting; it usually consists of two cells which open longitudinally to discharge the pollen, though in other plants there are various other ways for its escape. The pollen, or male element, consists of minute grains, usually rounded, and is generally yellow. The pistil consists of three parts: the lower distended portion (often more distinct than in this plant), the ovary; an elongated portion, the style; and at the apex a part without epidermis, and usually viscid, the stigma. The ovary, being cut open longitudinally, is found to contain (in this case) numerous small pulpy bodies, the ovules.
The influence of the pollen falling upon the stigma causes a growth to take place within these ovules, which results in the formation of seed. Such being the parts of the flower, and such their general functions (to be more fully mentioned further on), it remains to point out the leading modifications these parts undergo to produce the great diversity flowers everywhere present. The pedicel or flower stalk and the floral leaves or bracts, not strictly belonging to the flower, are placed so near it that they may affect its general appearance, or that of the flower cluster taken as a whole; in the Venetian sumach or smoke tree, the flowers are very small, but the pedicels, especially those which bear no flowers, are thickly clothed with hairs and increase in size, so that the conspicuous part of the inflorescence is these plumed stalks. The bract, besides differing in size and form from ordinary leaves, in many plants becomes petal-like and conspicuous; in the linden or basswood, each peduncle, or common flower stalk, has at its base a long ribbon-like bract attached for about half its length to the peduncle, and of a pale straw color which makes it nearly as conspicuous as the flowers. (See Linden.) In some sages, especially salvia splendens, the bracts are larger than and quite as brilliant as the flowers themselves, and much of the showy character of the plant depends upon them; while in euphorbiapulclierrima (best known as Poinsettia) the plant is cultivated solely for its gorgeous scarlet bracts, which surround a cluster of inconspicuous flowers The appearance of a whorl of bracts as an involucre was noted in describing the umbel.
Flowers of the largo family of composite have involucres often of many series of closely overlapping bracts (e. g., thistles and sunflowers), in some cases leafy, and in others so unlike leaves as to be reduced to mere papery scales. The terurination of the flower stalk, already described as the receptacle, and also called torus, is ordinarily slightly enlarged to make room for the parts of the flower which are attached to it; in some cases it is prolonged within the flower, as in the geranium and magnolia; in the passion flower it elongates and lifts the pistil far above the floral envelopes, and this occurs in other flowers; in the strawberry it is short and conical to accommodate the numerous pistils, and after flowering becomes large and juicy, and is the edible portion, while the proper fruits are the little seed-like bodies upon its surface. The receptacle in the rose, instead of bearing the pistils on its outer surface, is hollowed out, and they are attached to the interior of this cavity, the rose fruit or hip, being in structure like an inverted strawberry. In the great water lily (nelumbium), the pistils are immersed in pits in the surface of a broad receptacle, which in fruit is several inches across.
In these examples the receptacle has been that of the single flower; the same name is also given to the end of the stem when enlarged to receive several flowers, as in the composite. Sometimes, as in Dorstenia, the receptacle bears the flowers sunken in cavities in its substance, or, as in the fig, the common receptacle is hollowed and has its inner surface studded with minute flowers; but these forms of the receptacle belong to the inflorescence rather than to the flower. The disk or torus, so conspicuous in the mignonette, sumach, and other flowers, sometimes adherent to the ovary and calyx, is a development of the receptacle. - Floral Envelopes. In the flower taken as an example the parts of the calyx and those of the corolla are all alike, and the flower is regular; both series of floral envelopes being present, it is complete; and the parts in each series being five in number, or in the stamens twice five, the flower is symmetrical; in descriptive, botany the opposite of these conditions is expressed by the usual prefix, as irregular, incomplete, etc. The floral envelopes collectively, whether of two series or only one, are termed the perianth.
If one of the sepals or one of the petals is unlike the rest in size or form, the flower becomes irregular; the violet has three of its petals larger than the other two, and the irregularity is increased by the prolongation of one petal into a spur. In the larkspur the calyx is petal-like and much larger than the corolla; one sepal bears a long spur, which gives the plant its name, while the petals, much smaller and of irregular shapes, are only four. In this case the symmetry of the flower is broken up, one petal being lacking, and this absence or suppression of petals may include more than one, even the whole series, when the flower is apetalous. If but one series of floral envelopes is present, it is always regarded as the calyx, even if, as is often the case, it is petal-like. Many families of plants are constantly apetalous, and it is not unusual to find apetalous genera in families where the majority of the plants have petals. Other variations are produced by the cohesion of the members of the same series; the sepals unite by their edges to form a monosepalous or gamosepalous calyx, and the petals to form a monopetalous or ga-mopetalous corolla; this cohesion varies greatly in degree, from being confined to the base of the parts up to their union into a complete tube, with every intermediate gradation.
Where the union is more complete with some of the sepals or petals than the others, as in a five-parted flower, where two are united in one body and three in another, a two-lipped calyx or corolla is the result. The united portion of a calyx or corolla of this kind is its tube, and the free portion its limb. The terms used in describing the forms of these are usually derived from common objects, such as bell-shaped, funnel-shaped, wheel-shaped, and others; while special names are given to other forms, as labiate to the ordinary two-lipped corolla, which is itself capable of a great number of forms, in some of which one of the lips is nearly obsolete; a two-lipped corolla with its throat closed by a projection, the palate, is called personate. In the composite ligulate or strap-shaped flowers are common, such as the rays of the sunflower; in this the union of the petals is complete except at one side, which allows the whole to lie flat, though the union is generally manifest at the base. A handful of wild flowers or of single ones from the garden will afford numerous illustrations of these modifications of the floral envelopes, and many others.
The office of the floral envelopes, the calyx and corolla, being to protect the essential organs, the stamens and pistils, we in most cases find one or both present; there are naked or achlamydeous flowers in which both are wanting, but here their place is generally supplied by a bract, which serves to protect the essential organs; the graceful and fragrant liz-ard's-tail-of our streams (saururus) affords an interesting example of naked flowers. - Floral Organs. The stamens (collectively the andrw-cium) are next within the corolla, and present a great variety in form and number; their filament or stalk may be very long, or so short as to be imperceptible; the anther usually consists of two cells, and these are frequently placed one at each side of the filament at its apex; the portion of filament between the cells is the connective, and this may be prolonged more or less beyond the anther, and assume various shapes; in the oleander it appears as a long hairy tail; in some instances it is widened in such a manner as to separate the two cells far apart; the filament is often broad and flat, and sometimes bears appendages at its base. In the common pansy and other violets it is on two of the stamens prolonged into a process at the back of the anther much longer than the filament itself.
In methods of attachment to the filament the anther presents some variety; it may bo at the very apex (innate), or with one cell at each side of the filament (adnate), or it may open toward the pistil (in-trorse), or away from it (extrorse), and again it may, as in the lily, be hung by the centre and move in any direction (versatile). The various shapes assumed by the two lobes are used in descriptive botany, as are the methods by which the pollen is liberated; instead of the usual longitudinal slit, each cell may have a pore at the top, or each may have, as in the barberry, one, or as in the sassafras and others of the laurel family, two valves, or trap doors, hinged at the top, which open to liberate the pollen. In respect to numbers, they range all the way from a solitary stamen to so many that the number is regarded as indefinite; when of the same number as the petals, they are usually alternate with them; when twice as many, then one is opposite the petal and the other alternate. The stamens sometimes appear in two or more definite series; often a certain number have no anthers and are abortive.
As with the sepals and petals, the stamens are often united; when their filaments cohere to form a tube (e. g., the mallow family), they are monadel-phous; if in two groups (e. g., pea), they are diadelphous; and they may unite in threes or larger clusters. Where the union takes place by the anthers, leaving the filaments free, the stamens are syngenesious. - When the anther has reached its full development it opens in one way or another for the exit of the pollen, which is usually a loose powder, most frequently yellow, but sometimes brown, purplish, bluish, etc.; usually globular or oval, it is sometimes polyhedral, in the pine of three, and in the cat-tail of four united grains. In many orchids it is bound together in pollinia by elastic threads (see Orchids), and in the milkweeds (Asclepias) it appears as solid waxy masses. In size the pollen grains vary from 1/10 millimetre to 1/100- or even less. Some kinds of pollen have a surface beautifully marked with spines, or by the unequal thickening of the membrane. In structure the grain consists of an outer (extine) and an inner (intine) membrane, and is within filled with a thick granular liquid, the fovilla, which is its essential part.
When pollen is moistened under the microscope, the grains swell and burst, either by slits or by pores, which are thinner parts of the outer membrane, and through these the inner membrane protrudes in what is called the pollen tube, which in turn soon bursts and sets free the fovilla. The number of openings in the outer membrane of the pollen grain varies from one to several, though but one is used. - The pistil (or gyncecium) occupies the centre of the flower, and to the nourishment, protection, and development of this all other parts are subservient. Its parts, ovary, style, and stigma, have been already pointed out. In the flower chosen for illustration there are five separate and distinct simple pistils, and these are arranged in a circle in the centre of the flower; often there is but one, which then occupies the centre, or if there are two, they stand face to face in the centre; its occasional elevation upon a prolonged receptacle has been referred to. In the pistil already illustrated the parts are not so distinct as in some others.
The ovary and stigma are the essential portions, and the style is frequently not manifest, the stigma being sessile directly upon the ovary (e. g., poppy), but in many cases it is much elongated, and as in colchicum several inches long, The pistil already referred to is what is called a simple pistil or carpel, having but one ovary, one style, and one stigma. As with other parts of the flower, the pistils sometimes cohere to form a compound pistil; this union may involve the ovaries only, or even only the lower parts of them, leaving the styles distinct; it may include the styles, the stigmas remaining separate; or all may be united throughout. The stigma, if terminal, may be a simple knob; it is sometimes lobed, and in some cases, as in Sarracenia, forms an umbrella-like body, with stigmatic points at the margin. (See Pitcher Plants.) In some flowers it is not conspicuous, but simply a space denuded-of epidermis; and it generally has a viscid secretion to hold the pollen which may fall upon it.
The ovary of a compound pistil, when transversely divided, usually shows as many cells as there are simple pistils entering into its composition; if the five simple pistils of our typical flower were to be consolidated into one, it would bring the placentae or ovule-bearing portions in the centre, and this position (axile) of the placentae is very common in compound pistils; but in some flowers (e. g., poppy) a compound pistil may be one-celled, with its placentae in lines upon the wall of the ovary, a peculiarity of structure to be hereafter explained. In the compound pistil formed by the union of simple ones, or carpels, the division walls of the cells, extending from the circumference to the centre, are structurally double, as may often be seen in the fruit. - The Ovule and Fertilization. A section of the simple pistil or carpel of the typical flower shows the ovules at one side; they are attached to two lines of loose tissue, the placentae, and in this case are numerous; often the ovule is solitary; sometimes there is a definite number, and again so many as to be indefinite.
It has been already intimated that the ovule is the undeveloped seed, but more strictly speaking it is the organ within which the seed .is to be formed; it at first appears as a minute papilla or pulpy swelling upon the placenta, the nucleus; soon a coating grows outside of and envelops this, which is soon followed by a second and similar coat, thus enclosing the nucleus within two sacs, having at the top corresponding openings, the foramen or micro-pyle; the ovule is supported upon a longer or shorter stalk, the funiculus. The ovule may be straight, bent over on itself, or bent down upon its long stalk so that it is completely inverted. When it is fully developed a cavity is formed in or near its centre, the embryonic sac, which is filled with mucilaginous material. If the ovule is not fertilized, i. e., does not receive the influence of the pollen, it with the ovary develops no further. If pollen falls upon the stigma the pollen tubes are protruded and make their way down through the loose tissue of the style, probably receiving nutriment from that to promote their growth, until the embryo is reached, where the act of fertilization takes place.
Physiologists are not agreed as to precisely in what the act of fertilization consists; the pollen tube has been traced from the stigma down through the tissues of the style to the foramen of the ovule; after this takes place there appears within the nucleus of the ovule an embryonic vesicle, constituting the commencement of a new plant, which begins with a single cell, formed through the influence of the pollen, and this by its increase through subdivision develops into an embryo or young plant, the coats of the ovule at the same time enlarging and changing to become the coats of the seed. But the changes in the embryo are not the only ones which follow fertilization; the ovary itself greatly enlarges and alters, and becomes the fruit, a name in botany applied to the ripened pistil and the parts attached to it, without reference to edible or other qualities. With the completion of the embryo the seed and fruit are mature. - Morphology of the Flower. Before describing the fruit, reference must be made to other modifications in the flower as well as to the nature of the flower itself.
The adhesion of parts of the same series has been described, but there is another union of parts which not only greatly modifies the flower and obscures its structure, but which often has a marked influence upon the character of the fruit. This is the cohesion of parts of different series: the sepals and petals may not only unite among themselves, but the calyx may unite with the corolla for a portion of their length; this union sometimes includes the stamens (e. g., peach and cherry), in which case the petals and stamens appear to be inserted upon the margin of the calyx tube, though structurally all are regarded as having their origin on the receptacle, and cohering for a part of their length. This union may proceed still further, and in part or in whole involve the ovary (e. g., apple and pear), when the other parts of the flower appear as if inserted upon the top of the ovary, giving rise to the term inferior ovary of the older botanists. It sometimes occurs that the calyx and corolla remain distinct, while the stamens and pistils are blended in one mass (see Orchids); such flowers are said to be gynan-drous. In describing the organs of vegetation some of the many forms assumed by root, stem, and leaf have been pointed out, and it has been shown that these are often turned aside from their ordinary uses and made to serve some eutirely different one.
Now when the plant is ready to provide for its reproduction by seed, is an entirely new set of organs called into existence, or are those with which the plant is already provided made to serve for this purpose also? What has been shown of the morphology of the organs of vegetation has prepared the way for the statement that a flower is only a peculiarly developed branch, and that its parts are only modified leaves. The theory of metamorphosis, as it is termed, does not assume that the parts of the flower were formed first as leaves and then transformed into sepals, petals, etc, but that they are homologous with leaves; and that plasticity which allows a leaf to appear at one time as a tendril, at another as a spine, again as a pitcher, or as a most efficient trap for catching insects, will allow it, when the plant requires such organs as stamens and pistils to produce seed, and calyx and corolla to aid in the work, to assume these forms, which in many cases are not further removed from the normal condition than occurs when there is no doubt that the organ is still a leaf. But a few of the many illustrations in support of this theory can be cited.
It has been shown that flower buds, and consequently flowers, appear in the same place as do leaf buds - terminating the principal stem, or sessile, or on small branches in the axils of its leaves; also that when a flower cluster is formed it results from a modification of the usual manner of branching. The flower terminates the stem upon which it is placed, whether the main stem or flower stalk, the usually enlarged and rounded end of the stem becoming the receptacle, upon which the parts of the flower are placed, one above another; in viewing a flower as a modified branch, we find its internodes very short, but scarcely shorter than in the houseleek, where they are so abbreviated as to bring the leaves so close together that they touch; indeed, except in color, some plants of that family closely resemble the flower of a double camellia or rose. Abnormal developments often throw much light upon the real nature of the flower and its parts, in which, the vital forces of the plant being from some cause disturbed, there is a reversion of the parts to the leaf or toward it, or retrograde metamorphosis as it is termed.
Cases have been already cited in which the receptacle is prolonged in the centre of the flower to make room for the numerous pistils; but it not rarely happens, in the rose, for example, that the receptacle is abnormally prolonged and assumes the true character of a branch, growing several inches in length, bearing true leaves, and being terminated by an another flower bud. The writer had a specimen in which this was repeated a third time on the same flower. The calyx is so generally green that its leafy nature is manifest, and often in the rose the divisions of the calyx become pinnate, like the ordinary leaves. The resemblance to green leaves is less manifest in the petals, as their more delicate texture and often brilliant colors aid in disguising them; but these colors and texture are often quite as striking in bracts, which are unmistakably leaves and in no wise attached to the flower. In many cases it is impossible to distinguish between calyx and corolla; where there are many series of these parts, as in calycanthus, it is very difficult to say which is calyx and which corolla; some of the double camellias of the greenhouse show this in a striking manner, and in the magnolia calyx and corolla are only to be distinguished by position.
In the green rose the place of the petals is occupied by a cluster of green leaves. In many flowers the petal is narrowed at the base into a claw, which corresponds to the petiole of the leaf, and this is sometimes, as in the pink, very conspicuous; petals with a manifest claw correspond to petioled, and those without it to sessile leaves. The stamen shows much less resemblance to the leaf than do the sepal and petal. The filament represents the petiole or stalk of the leaf, and the theoretical structure of the anther is that it corresponds to the blade of the leaf with its edges curved in toward the midrib to form the two cells of which most anthers consist; the midrib of the leaf is the connective; the cellular tissue of the leaf undergoes marked changes and is developed as pollen. The stamens are frequently transformed into petals, which are undoubted leaflike organs, as may be seen in any partly double rose and many other semi-double flowers; if stamens are visible in a double rose, it is generally easy to find bodies in every intermediate state, from a nearly perfect petal with the vestiges of an anther upon its edge, a body half petal and half stamen, to a nearly perfect stamen with a fragment of petal developed upon it.
The double columbine of the gardens affords a still more striking illustration; in this flower the petals are in the form of a hollow curved spur, and the stamens may be found with a filament, but bearing in place of the anther one of these curved petals. The rose often shows a complete series of transformation from the calyx to the stamen: first perfect sepals, within these petals with a green midrib or partially changed to sepals, then perfect petals, and within these the intermediate states between petal and stamen. The double rose is a flower in an abnormal state, but we find similar gradations in wild flowers unchanged by cultivation; a striking example of this is shown in the white water lily (nymphcea), in which the calyx is green on the outside and petal-like within; within this are several series of petals, some of the inner ones with an abortive anther at the tip, and so on toward the centre, the petal gradually disappearing and the anther more perfectly developed, until true stamens are found bearing no resemblance to petals. - In some compound pistils it is difficult to trace any resemblance to the leaf, but when the pistils or carpels are distinct, especially in flowers with a solitary pistil, the foliaceous character is manifest.
It is not difficult to see that the pistil of our typical flower, or that of the larkspur, peeony, or the common pea, may be formed by folding the edges of a leaf together and uniting them to form a hollow sac or pouch; that this is the real nature of the simple pistil may often be seen in the double cherry, in which the retrograde metamorphosis is strikingly exhibited, an infolded leaf actually standing in place of the pistil, its prolonged midrib, the style, bearing the knob-like stigma at the end. Other plants afford illustrations of the same point, especially an alpine strawberry, which produces all parts of the flower, even the stamens, as green foliaceous bodies, and the pistil, though minute, as a bud of overlapping leaves. Taking this view of the simple pistil, that it is an infolded carpel-lary leaf, several theories have been proposed as to the nature of the ovule, and the subject has given rise to much discussion. Schleiden regards the ovules as buds produced upon the edges of the carpellary leaf, and the placenta as a cellular growth from its margins; though the edge of a leaf is an unusual place for buds to appear, some plants naturally produce them there, especially oryophyllum, which propagates itself from buds upon the leaf.
This view of the homology of the ovule is supported by the fact that a number of abnormal pistils have been observed in which leaf buds appear in the place of ovules; a case of this kind is a monstrous columbine, in which the simple pistils develop as flat leaves, and bear leaf buds upon their edges. Accepting this view of the structure of the simple pistil or carpel, that part of it which corresponds to the midrib of the leaf is the dorsal suture, and the union of the edges of the leaf forms the ventral suture, the placenta being always borne here. When two or more simple pistils are united, it is by their ventral sutures, and the collective placentae will be in the centre; the placenta is sometimes free in the axis of the compound ovary, a state of things often caused by the obliteration of the dissepiments or division walls. In many compound ovaries the placentas are parietal, or upon the sides of the ovary; a pistil of this kind is regarded as composed of several carpellary leaves, which, instead of infolding and uniting in the manner already described, are placed in a circle and joined by their contiguous edges; the placenta in this case, instead of being composed of two edges of one carpellary leaf, is formed by the edges of two different adjacent leaves.
By the projection of the parietal placentas toward the centre of the ovary a great variety of forms are produced, some of which are quite puzzling. - Double Flowers. These, to which frequent reference has been made, rarely appear in nature, though those having normally more than one series of petals may sometimes be taken for such. In horticulture a flower is regarded as double when its proper number of petals is much increased; in many cases the beauty of the flower is greatly enhanced by doubling, as in the rose; but those flowers the beauty of which is largely dependent upon the regularity and symmetry of their parts, the lily for example, are only disfigured by becoming double. Doubling is most frequently produced by the conversion of the stamens into petals; but in many cases it is due to the production of two or more petals in the place of one, and there are double flowers which neither of these causes fully accounts for. Doubleness is regarded as indicating a tendency to sterility, and whatever disturbs the reproductive functions tends to produce double flowers; fertility is diminished by a variety of causes, and long cultivation in a rich soil is one of the most common.
When any derangement of the forces of the plant causes the stamens to be sterile, they revert to petals; when this once takes place, the flower "breaks," as the gardeners say, the tendency becoming hereditary; seedlings from a plant thus changed are in some cases likely to show a greater departure from the normal condition, and by continuously selecting seeds from those in which the tendency to become double is strongest, the habit may be fixed; finely doubled flowers have been produced in this manner from one in which only a single extra petal appeared. The stamens become sterile first, and a flower may have all of its stamens converted into petals, while the pistil is still fertile; such flowers fertilized by the pollen from a single or partially double one will yield seeds with a strong tendency to produce double flowers. When the metamorphosis is carried so far as to involve the pistils, the plant can only be propagated by cuttings or other subdivision. The doubling in flowers of composites is not due to a change in the stamens; in the natural state of the dahlia, sunflower, and others, there is a disk of small tubular flowers, surrounded by a ray of much larger flat and showy ones; these flowers are regarded as double when the tube-shaped corollas of the disk are developed like those of the ray.
The Poinsettia has been mentioned as a plant cultivated for the showy bracts which surround a cluster of small inelegant flowers; a double variety of this was discovered by Roezel in Mexico; in this the flowers are changed into leafy branches, and these leaves are colored like the ordinary large bracts, exhibiting a large mass of brilliant color without any flower at all being concerned. - The Fruit. This has already been described as the ripened (simple or compound) pistil, and such accessory organs as remain attached to it. The changes the pistil undergoes in maturing are often very striking; one of these is a great increase in size; another frequent change is in the tissues, which may become exceedingly pulpy and soft, or on the other hand may be indurated and even become bony. Another marked change occurs in the substance; the pistil, which at first was essentially the same in composition as a leaf, as it enlarges becomes, as in the grape, charged with acid, or in the persimmon with tannin, and as the fruits reach maturity the acidity or astringency gives place to sweetness, indicating the formation of sugar.
The interior structure of the pistil is sometimes not to be traced in the fruit; the number of cells may be diminished, or by the formation of false partitions increased; the number of seeds perfected is often fewer than the ovules; while the peach, almond, and their allies always have two ovules in the ovary, but one generally becomes a seed, though both sometimes develop, as " double-meated " almonds and peach stones are not rare. In the oak the ovary has three cells with two ovules in each cell; in ripening the three divisions are obliterated, and five ovules perish, as we find. the acorn to be only a one-celled, one-seeded fruit. A few examples drawn from familiar plants will illustrate the changes the pistil undergoes in becoming a fruit. One of the important characters in descriptive botany is the dehiscence, or way in which the fruit opens; many fruits, both simple and compound, are indehiscent, but in other cases fruits of both kinds open to let out the seed; the dehiscence in a simple carpel may be along the ventral suture, or by both sutures; and in a compound fruit the carpels may open upon the back, or they may break away from one another and open by the ventral suture; there are several anomalous forms of dehiscence which seem to bear no relation to the structure of the pistil, as where the fruit opens by a regular lid.
The ripened ovary, or the wall of the fruit whatever changes it may undergo, is the pericarp. In the common buttercup the fruit is small, dry, one-seeded, and indehiscent; such fruits are akenes; the proper fruit of the strawberry is of this kind, the edible portion being the enlarged receptacle. The fruit of the columbine, psaony, and larkspur results from simple pistils; these open at the ventral suture and expose the numerous seeds; such a fruit is a follicle, from which to the legume or pod is but a step; in the pea it is dehiscent by both sutures, and the carpel falls apart in two valves; in the pea the pericarp still retains something of its leafy character, and in the common bladder-nut the walls become very thin and papery, and before they are fully matured are strikingly leaf-like. The most remarkable change in a simple pistil is seen in the drupe, of which the almond, peach, plum, cherry, and others known as stone fruits, are examples; in these the car-pellary leaf undergoes two very unlike changes; the interior portion of the ovary, that corresponding to the upper part of the leaf, as the fruit matures has its cells filled up by a deposit which makes them nearly solid, and the tissue which they form, the stone of the fruit, exceedingly hard and bony; while this is going on the outer portion, or the under side of the leaf, increases greatly in thickness and becomes fleshy, juicy, and edible, as in the peach and others, while in the almond, though it becomes thick, it is hard and dry.
The stone (puta-men) in these fruits is not a part of the seed, but of the pericarp. Of the same structure with these conspicuous examples of the drupe is the very small drupe of the raspberry and blackberry; in these each ovary becomes a minute stone fruit; these as they grow are much crowded, and in the raspberry they cohere, but when mature they separate entirely from the receptacle, which has nourished them, as a little cap; in the blackberry the little drupes retain their hold upon the receptacle, which becomes more or less juicy and edible. Berry is a general name for fleshy fruits, without reference to the number of carpels; in the papaw the fruit is an elongated very pulpy berry, resulting from a simple pistil; in the grape the pistil has two carpels, and in the gooseberry and tomato three; in the tomato cultivation has brought the fruit into an abnormal condition, and while its botanical structure is broken up and confused, it is much improved in an economical point of view, as the fleshiness in this belongs mainly to the placenta.
A pepo is a berry with a hard rind, of which the squash, melon, and all of the gourd family are illustrations; in the proper gourd the rind when ripe is very hard and woody, and the pulp dry; in the watermelon and cucumber the rind never becomes very hard, while the pulp is very juicy. In the orange, lemon, and others of the family, the fruit results from a many-celled ovary; when ripe it has a leathery and often very thick. pericarp; the cell walls remain and divide the fruit into sections, which are filled with separable cells containing a watery juice; this kind of fruit is called a hesperidium. Of dehiscent fruits, or capsules resulting from a compound pistil, the crucifers or mustard family present a simple form; the mustard, cabbage, and others have the form called si-lique, in which the ovary consists of two carpels and has two parietal placentae; the capsule usually falls apart in two valves, leaving the framework to which the seeds are attached. In the iris, the three carpels break open on the back (loculicidal dehiscence), while in the col-chicum the cells open at the ventral suture (septicidal). The poppy capsule liberates its seeds through a number of openings below the persistent stigma; in the common purslane of the gardens the capsule opens by a transverse fissure, the upper half falling away.
In inde-hiscent fruits the pericarp sometimes decays to set free the seeds, and in other cases it is ruptured by the swelling of the contained seed when it germinates. Among fruits formed by the aid of other parts than the ovary, the most common is the pome, of which the apple is a familiar illustration; as in the flower of the apple the calyx is so united with the ovary that it appears as if seated upon it, so in the fruit we find this union of calyx and ovary still continued; in the apple the proper carpels are the parchment-like bodies which are found at the core, and when the fruit is cut across appear arranged in a circle in the centre, and enclosing the seeds; these are immediately surrounded by a pulpy expansion of the receptacle, and outside of the whole is the enlarged succulent calyx tube which makes up the greater bulk of the fruit, its persistent free divisions generally remaining in a withered state when the apple is mature. In the aromatic wintergreen or checkerberry, the edible portion of the fruit is the enlarged calyx, not in this case united with the ovary, which is a dry capsule loosely invested by the pulpy calyx.
The grain (caryopsis), of which the cereals are examples, results from a one-seeded ovary, the seed usually adherent to the proper pericarp, which becomes very thin and papery, and is commonly known as the hull. Fruits which result from several distinct flowers closely crowded upon a common axis are aggregate fruits, of which the pineapple is a remarkable illustration. (See Pineapple.) The mulberry shows a similar structure on a much smaller scale. In the pine family the pistil is an open carpellary leaf or scale, and the same structure is retained in the fruit, where the scales enlarge and become woody, as in the pine and related genera (see Pine), or they become fleshy, as in the juniper and some others. The cashew (anacardium), a tropical tree, presents a most anomalous fruit, it being no part of the flower; the pistil ripens into a nut, sometimes found in commerce, the cashew nut, and at the same time the peduncle or flower stalk undergoes a change, swelling into a pear-shaped, pulpy, edible mass called the cashew apple, upon the top of which the nut rests. - The Seed. The proper nourishment, maturing, protection, and final distribution of seeds are the ultimate object of all the varied forms of the fruit. The internal structure of dicotyledonous seeds has been sufficiently described.
Monocotyledonous seeds present an equal variety in their structure and methods of germination. In some cases the embryo is an elongated cylindrical body, in which no distinction of parts is apparent until germination takes place; the lower end of an embryo of this kind (e. g., onion) is the radicle, and the upper portion is a cotyledon, wrapped around a plumule, which shows itself when the seed germinates; on the other hand, maize or Indian corn has a highly developed embryo, in which the radicle is distinct, bearing a single large cotyledon partly enveloping that and the plumule, which consists of several rudimentary leaves closely enwrapping one another. In monocotyledonous plants the growth of the primary root soon ceases (see Palm), abundant secondary roots being formed from the lower nodes of the proper stem or axis, as may be seen in young plants of maize. - The ovule has been described as having two coats; these when the embryo is ripe become the seed coats; the inner, the tegmen, is sometimes manifest, but it is often united with the outer or wanting altogether. The outer seed coat, the testa, varies greatly, and it is due to the character and markings of this that seeds differ so greatly in their external appearance.
It may be soft and membranous or hard and bony; its surface is often marked by furrows, pits, and other irregularities; it sometimes expands into a broad wing (e. g., catalpa), or produces a tuft of silky hairs (e. g., milkweed). The appendages to the seed coat of one plant are of vast importance to the world; the fibre, cotton, is only elongated simple cells produced from the spongy testa of the seed coat of gossypium. These and other appendages aid in the dispersion of the seeds; the one-seeded akenes of the composite family, often mistaken for seeds, also present a wonderful variety of contrivances for dissemination. The stalk of the ovary (funiculus) retains the same name in the seed. In falling away from the stalk when ripe the seed shows a scar upon its testa, the Mlum, sometimes a mere point and obscure, often quite noticeable, as in the bean, and in the horse chestnut very broad and conspicuous. Some appendages of the seed are due to an expansion of the funiculus, while others are regarded as a growth from the micropyle, but the distinction between the two is not always manifest.
The seed of the white water lily (nymphcea) is surrounded by a delicate transparent sac (arillus) of this nature; the pulpy covering of the seeds of the burning-bush (euonymus) and waxwork (celastrus) is an aril, and in the nutmeg the same organ becomes an important product, mace. The content of the seed, whether it consists of the embryo only, or both that and the albumen, is the nucleus, the structure of which has been sufficiently described in speaking of germination. - In this sketch of the plant but little reference has been made to the great series of cryptogamous or flower-less plants, which are subdivided into several classes, each having its peculiar structure and method of reproduction; accounts of these may be found in the articles Alg - , Ferns, Fungi, Liverworts, Lycopodium, and Mosses. - Classification of Plants. While plants everywhere are engaged in essentially the same work, that of vegetation or the growth of the individual, and reproduction or providing for •the continuation of the kind by seeds, the details of these operations, as has been shown, are wonderfully varied; to study such a multitude of plants understandingly, they must be grouped and arranged into a system.
Some account of the different methods that have been from time to time employed is given in the article Botany. The natural system, attributed to Jussieu, who more distinctly presented it than any of his predecessors, has been greatly modified and improved by the labors • of subsequent botanists; the most recent presentation of this is in the " Synopsis " appended by Dr. J. D. Hooker to the-English edition of Le Maout and Decaisne's " General System of Botany," translated by Mrs. Hooker; in this the arrangement to be adopted in Hooker and Bentham's Genera Plantarum is indicated. Jt has already been shown that flowering plants are divided into two great classes founded upon the structure of the seed, monocotyledons and dicotyledons, which are accompanied by differences in the structure of the stem, called endogenous and exogenous growth. These are subdivided into subclasses, divisions, and other groups founded upon the structure of the flower. Classification by whatever system commences with grouping individuals into species; it has been suggested that it is difficult to define the limits of the individual in plants, as they are made up of a multitude of parts, each of which with proper care may be separated and continue an independent existence.
If the question of individuality has given rise to much discussion, not less so has that of the species. It is not easy to give a brief and satisfactory answer to the question, What constitutes a species? One of the best definitions is, that it is a collection of individuals so nearly alike that they may have been the progeny of one parent. Some botanists believe that a species may varv widely, and others rank what the former would consider mere varieties as distinct species; while Bentham considers that the various forms of the European blackberry and dewberry are reducible to two species, some other botanists enumerate 36; the rich collection of facts brought together by Mr. Darwin in his " Origin of Species" and "Variation of Plants and Animals under Domestication " shows the wide variations of which species are capable, and has led naturalists to take very liberal views of their limits. This liability to vary, and the fact that peculiarities in species may be rendered permanent, are of the greatest importance economically; while many desirable departures from the normal form can only be continued by subdivision, others, as in annual and biennial plants, may be propagated by the seed; by careful selection of plants as seed-bearers, and continuous sowing of seed for several years in succession, a peculiarity becomes fixed and a race is produced.
Species are grouped into genera founded upon similarities of structure, and where the genus is a large one it is divided into subgenera, and these again into sections. We find genera large or small, according to the importance attached to points of structure by different botanists; while most regard the pine, spruce, larch, and others as sufficiently unlike to rank botanically, as they do popularly, as distinct genera, some class them all in the pine genus. Kindred genera are grouped as families or orders; these when large are often subdivided, and we have suborders, tribes, and subtribes. The peach, strawberry, blackberry, rose, and apple have certain characters in' common, which bring them all into the rose family; but they differ so much in other respects that they fall into three subfamilies, the peach in one, the apple in another, and the others in a third; still further, the strawberry, blackberry, and rose, though in one subfamily, are sufficiently unlike to be placed in separate tribes. The next group above orders is usually the division, of which in dicotyledonous plants there are three, viz., the polypetalous, monopetalous, and apet-alous divisions; and in monocotyledons there are similar divisions founded on the relations of the floral envelopes to the ovary.
Within these divisions the families are grouped in series and cohorts or alliances, bringing together those most nearly related in structure. - Plant Distribution. The distribution of plants upon the surface of the globe (geographical botany) forms a distinct and important department of botanical science, which can be merely glanced at here. The study of the distribution of plants as we now find them involves not only a knowledge of the requirements of each species, but of the geological and geographical changes which preceded the present epoch, and the various natural and artificial agents which have affected the distribution. In treating of germination it has been stated that while seeds will vegetate at a quite low temperature, and may endure a very high one, yet there is a certain point most favorable to each kind of seed. It is so throughout the whole life of the plant; a species may maintain its existence in an unnaturally low or high temperature, yet there is a degree of heat most favorable for each to perform its functions of vegetation and seed production; hence temperature is one of the chief agents in determining the distribution of plants.
It is a well known fact in physical geography that latitude alone does not govern the temperature of a locality, but that altitude must be taken into account; on the Alps the harvest is 17 days later for each 1,000 ft. of altitude, and a similar rule holds elsewhere; it is a matter of common observation that as we go south in our own country, if we would find the common plants of the northern states in Georgia or North Carolina, we must seek them in the mountains. In general terms it may be said that each species requires a certain number of days to complete its career of vegetation and reproduction, and that during these days it requires a certain mean temperature; the mean temperature multiplied by the number of days gives the sum of heat the plant demands for its development; within certain limits, if the temperature be lowered, the time must be prolonged, or if the mean temperature of the days is higher, their number may be lessened; hence we find vegetation in high latitudes often remarkably rapid, and though the south of England is nearly 10° south of Upsal, Sweden, the wheat harvest in both localities takes place at the same time.
As a general rule a plant will flourish, so far as temperature is concerned, wherever the sum of heat is sufficient for its needs; it is not necessary to state here the exceptions to this. Another important condition influencing plant distribution is moisture; the plants native to arid regions, such as the elevated plains of Arizona and New Mexico, are especially adapted in their structure to resist a drought of many months, and would die if taken to regions of frequent rains, as quickly as would plants from moister localities transferred to these plains. Certain plants that live only under the shade of trees are not found in treeless regions, nor are certain others met with away from the influence of the sea. Peculiarities of soil have also their influence on distribution, and there is as much difference between the plants of sandy and heavy soils and of calcareous and granitic ones as between those of swamps and dry hilltops. Some naturalists hold that all the plants of a particular species, as at present recognized, are descended from a single parent individual created " in the beginning;" others that there were several distinct but widely separated "centres of creation;" and others, with Darwin, that species as we now know them were derived from other and simpler " primordial forms." But whatever may have been- the " origin of species" now upon the globe, the fact must be admitted that they have been transferred from the localities they occupied at first and become naturalized in others, for the distribution of plants is still going on.
While according to De Candolle no single flowering plant is really a cosmopolite, he enumerates 117 which extend over more than one third of the earth's surface, and 18 which are found over one half of the globe. In contrast with those widely distributed species, there are many of very limited area; the Kerguelen cabbage, found only on the small island which gives its name, is an instance of this; and again plants of the same kind are found in localities so widely separated as to make it difficult to trace them to a common origin: schizcea pusilla, a very local fern in the pine barrens of New Jersey, was thought to be peculiar to them until it was found in New Zealand; and more than one of our rare plants in the northern states is found wild elsewhere only in the Himalayas or in Japan. Many of the natural methods of disseminating plants are now at work, and it is inferred that they were in operation in early times. The seeds of some plants are carried to long distances by ocean currents, though the number of seeds which can resist the action of sea water is found to be comparatively small; rivers do much more in the way of distributing seeds, as do casual mountain torrents; birds may carry aquatic plants long distances, attached to their feet, and birds of passage perform an important part in dissemination, voiding the undigested seeds at a great distance from the point where the fruit was eaten; quadrupeds do their share of the work, and many seeds and fruits are provided with hooks which allow them to be attached to the hair and skin of these animals.
Man performs his part in the distribution; many plants (especially in the tropics) have escaped from cultivation and become so thoroughly naturalized as to appear as natives, and the weeds of agriculture are to be found in every settlement however remote. "Wars have had their effect upon vegetation, and the passage of an army through a country leaves weeds in its train which were not there before. After our civil war a little leguminous plant, Les-pedeza striata, sprang up all over the southern states; it is not known how it came or where from, but its native country is Japan, and in some localities as " Japan clover " it is valued as a forage plant. In a geographical classification, plants may be arranged in zones as regards latitudes, or in zones as to altitudes; or following Schouw, the globe may be divided into phyto-geographic regions founded upon the prevailing plants, such as the region of mosses and saxifrages, the region of palms and melastomas, etc. Besides these there are vast numbers of marine plants the distribution of which is marked by distinct bounds.
The accompanying chart is designed to present at one view the prevailing vegetation of the different parts of the globe, and to give an idea of the geographical limits of the principal cultivated and economical plants. - Bibliography. In this sketch of the plant, only the leading points have been briefly treated, and some are necessarily omitted altogether. Naturalists are now much occupied in studying the relations of plants and insects, and the reader is referred for some account of these to the titles Insect Fertilization and Insectivorous Plants. Since the last named article was written, Sir John Lubbock's work, "British Wild Flowers considered in Belation to Insects" (London, 1875), has appeared, and gives many new observations. To aid those who would follow the subject of this article more in full, the following works are enumerated, as supplementary to the more extended list (mainly of systematic works) given under Botany. Plant structure and plant life are presented in a form attractive to young people by Prof. Asa Gray in " How Plants Grow" and "How Plants Behave" (New York, 1858 and 1872), and by Miss Eliza Youmans in her "First" and "Second Book of Botany" (New York, 1870-'73). The general reader will find Gray's " Lessons " (1868) sufficiently comprehensive, and his " Structural and Systematic Botany" is a thorough work suited to advanced students.
The elaborate work of Le Maout and Decaisne, translated by Mrs. Hooker and edited by Dr. J. D. Hooker under the title " A General System of Botany, Descriptive and Analytical" (1873), has been referred to as the most recent work on classification; it is prefaced by a full and profusely illustrated treatise on structure. The student who would study the physiology of plants and their relation to the soil will find in Prof. S. W. Johnson's " How Crops Grow " and " How Crops Feed" (New York, 1868 and 1870) the most complete presentation of the subject. The literature has recently been enriched by a translation of Julius Sachs's Lehr-ouch der Botanik, with the title " Text Book of Botany, Morphological and Physiological," by A. TV. Bennett (London, 1875). The most complete work on the distribution of plants is A. de Candolle's Geographie botanique raison-nee (Paris, 1855). Darwin's " Origin of Species," "Variation of Plants and Animals under Domestication," "Fertilizationof Orchids," and " Movements of Climbing Plants" are full of interest to the general reader and wonderful storehouses of facts for the student.


Fig. 1. - Protococcus.
Plant of a single cell, multiplying by subdivisions (magnified).

Fig. 2. - Plant of Elongated Colls, placed end to end (magnified).
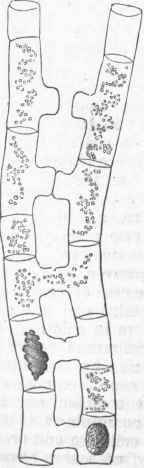
Fig. 8. - Conjugation in Zyg nema (magnified).
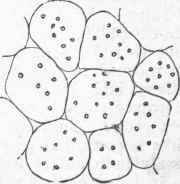
Fig. 4. - Cellular Tissue and Intercellular Spaces (magnified).

Fig. 5. - Woody Fibre and Intercellular Spaces (magnified).
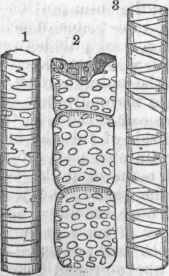

Figs. 6 and 7. - 'Vessels and Ducts from the Stem of a Melon. 1. Eeticulated. 2. Dotted. 3. Annular. 4. Spiral Vessels. 5. Dotted Fibre of Clematis (magnified).
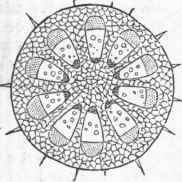
Fig. 8. - Section of Melon Stem. Bundles of Wood Fibre surrounded by Cellular Tissue.
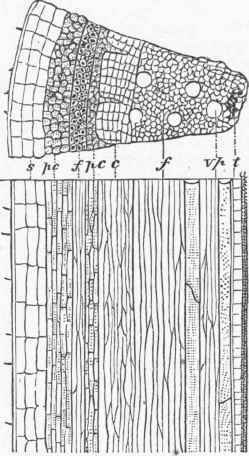
Fig. 9. - Maple Stem one year old. s to c, bark; c to t, wood; s, corky layer; pc, green layer; f, liber; c, cambium layer; f, woody fibre; vp, dotted ducts; t, spiral vessels.
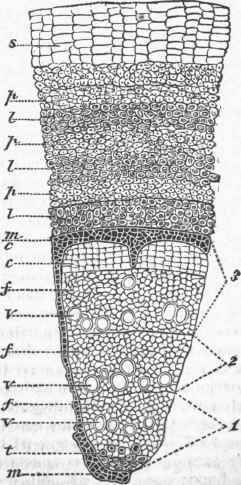
Fig. 10. - Maple Stem three years old. 1, 2, and 8, wood of first, second, and third years; c, cambium layer, beyond which is the bark of three years; mc, cellular tissue connecting by medullary rays with the pith m; I, liber. Other letters refer to the same parts as in fig. 9.
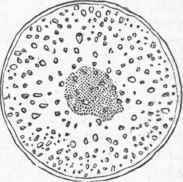
Fig. 11. - Section of a Mono-cotyledonous Stem (Indian Corn).
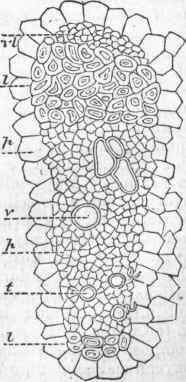
Fig. 12. - Bundle of Fibres from the above, containing the same parts as a dicotyledonous stem. (See fig. 10).
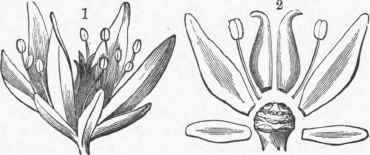
Fig. 13. - 1. A complete Flower of Sedum. 2. Dissection showing receptacle and two members of each, series, sepals, petals, stamens, and pistils.
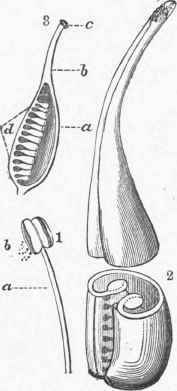
Fig. 14. - 1. Stamen: a, filament; b, anther discharging pollen. 2. Pistil and cross section. 3. Pistil cut lengthwise: a, ovary; b, style; c, stigma; d, ovules.
Continue to:
My Books

